Translocation of Organic Solutes:
Plants synthesize food materials by photosynthesis and store the same in the form of starch grains in the chloroplasts found in mesophyll cells. Then, they found starch is converted into simple sugars; then it is transported to regions, where it is required – stem apex, young flower buds, fruits and storage organs, which are the sites into which the organic food is translocated. The site of synthesis and the site of need are separated by time and space. The structures found between these sites play a significant role in translocation.

Potato plant with its well developed tubers in the soil.

Structures responsible for translocation;
Girdling experiments involving the removal of a ring of cortex including phloem cells below leaves prevent the flow of food materials towards roots which accumulate at the upper rim of the rig and shows swelling. Radioactive tracers like 14CO2 sucrose phosphates supplied are found to be translocated through sieve tube elements. While mealy bugs pierce their stylets into sieve tubes and suck the sap, the white bugs have specialized in feeding on the sieve tube sap. Thus, various studies reveal that the food material is translocated through sieve tubes.



Rate of translocation
Using radio active tracers at the source of supply like leaves, it is possible to measure the rate of translocation. Such experiments have shown that, the organic food material is translocated at the rate of 100-200 cm per hour which is relatively 1000 times faster than the rate of passive diffusion. However, not all plants exhibit the same rate of transport and it varies from plant to plant varietis; added to this, various factors also contribute to the rate of movement.
Direction of movement:
Food material always moves from the leaves of different regions of the plant body. Organic solutes found in the leaves nearer to the stem apex move upwards, and leaves found at the base move down wards. But the substances in the leaves found in between, move in both directions. But the bi-directional movement takes place in the same sieve tube or in different sieve cells is a debatable point. Some claim that it is unidirectional and others view it as bi-directional, that too in the same cell.

Translocating substances
Analysis of mealy bug’s stylet drops of sieve tube sap indicate that, the major amount of the sap is made up of sucrose. Though glucose and fructose are found in traces, they are not the tube translocates but they are the degraded products of sucrose in transit. Along with sucrose, rhamnose, and other sugars are also found to be translocated but in traces. Amino acids, other organic acids and phytohormones are also found to be translocated through sieve tubes.
Factors controlling the rate of translocation
1. Temperature: Organic solutes that move out of leaves towards their respective destinations are thermo-regulated. Increase in the temperature of soil increases the rate of movement of materials towards roots; decrease in the temperature results in the fall in the rate of movement of solutes in the leaves. Thus, the rate of metabolism of both supply end and receiving and have a direct relationship to the rate of translocation.
2. Light: Light is an important factor that affects the life of the plant in many respects like growth, flowering, photosynthesis, germination, etc. Besides these effects, is has a control over the translocation of food materials. Generally during day time, only a small amount of food materials is translocated out of the leaves. Out of this small amount, 2/3rd moves towards stem tip and the rest towards root. On the contrary, most of the food is translocated from the leaves during night or dark conditions. But more of food is translocated to roots than to stem at nights. The most favorable action spectrum has been found to be red light. Metabolic state of tissue: Actively dividing tissue needs more energy for its activities hence it needs more food materials; hence more of solutes move into these regions. Similarly, a winter bud, whose growth activity has come to a stand still, does not require more energy; so it requires very little amount of food. Thus, the metabolic status of the cells creates the demand and the supply has to be met with.
Another important aspect of the metabolism is the supply of energy rich molecules like ATP for translocation. Use of respiratory inhibitors like KCN, DNP etc., which inhibits the synthesis of ATP, have a drastic effect on the rate of translocation, there by it indicates that the translocation is an energy dependent process. Anatomical evidences show that companion cells present at the side of sieve tube cells are endowed with the cellular machinery to provide the needed energy for transport in the sieve cells..
Inorganic mineral deficiencies:
Inorganic nutrients are required for the normal functioning of the cellular processes, but the deficiencies of some inorganic ions, particularly born and phosphorus have a strong inhibitory effect on the rate of translocation. Boron has been found to be involved in the translocation. It not only enhances the translocation, but also facilitates the loading of the veins, how?.
Phytohomones
Growth regulating substances like auxins and cytokinins have a profound effect on cellular process like growth and differentiation. The exogenous supply of these enhances the rate of translocation. Whether they have a direct effect or indirect effect, is not clear. However, it is quite possible that these hormones activate the cells in the shoot and root spices, thus create a demand for the food. Thus, they may enhance the rate of translocation. As they also increase the rate of respiration, they may facilitate the increased rate of translocation.
Mechanism
While considering the mechanism of translocation, it is pertinent to consider various aspects of translocation and translocating structures. Translocating organs i.e., sieve tubes are living cells, the protoplasm shows all cellular activities including Protoplasm streaming. The longitudinal columns have sieve plates, through which plant proteins pass through and extend the entire length of cell forming a continuum between the cells. Companion cells are in close contact with sieve cells. Most of the energy demand of sieve cells is met with by companion cells which contain large number of mitochondria. In order to explain the translocating process, various mechanisms have been proposed and none of them satisfies all aspects of translocation. Historical account: From de Vries, time to the recent past, plant physiologists have come out with one or the other mechanisms like (1) Diffusion theory (2) Protoplasmic streaming theory (3) Transcellular theory (4) Pulsating theory (5) Mass flow theory.
In the case of diffusion theory, the main premise on which the explanation is sought is that solutes, like sucrose diffuse from the site of synthesis to the site of need. The diffusion has been considered as active diffusion. Whether it is passive or active, the rate of diffusion is many thousand times less than the rate of diffusion is many thousand times less than the rate of actual translocation. However, a diffusion gradient exists between the two ends of the translocating system.
Protoplasmic theory visualizes that protoplasm exhibits active streaming movement. As the protoplasm between the sieve tubes could perform bi-directional movement in the same cell, protoplasmic streaming can move the material enemas very fast (rate is not known). However, the rate of protoplasmic streaming is far below the rate of translocation observed. If protoplasmic streaming is the mechanism, then microfilaments which are known to be involved in this process should be present. What happens if the microfilaments are disrupted by inhibitors? Unfortunately, no such experiments have been conducted to prove their point.

Visualization of transcellular strands is another case of misunderstanding and misinterpretation. No one has observed the presence of such ‘dense’ transcellular strands.
Pulsating theory, which visualizes an active process, has some evidence because ATP supply is essential to maintain the observed rate of translocation. But now the pulsation is brought about, and what are the structural elements responsible for such pulsation have not been explained anywhere. Thus, the beginning this theory is also the end of it.

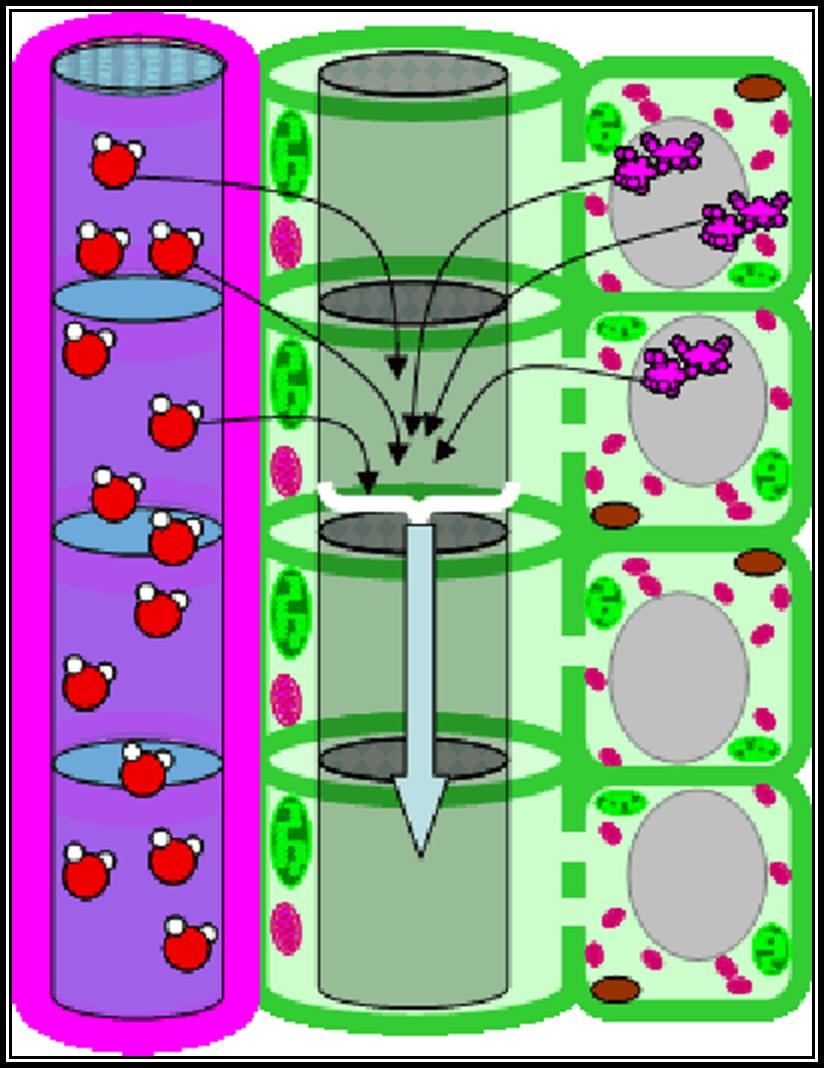
However, Munch’s Mass flow or pressure flow theory enjoyed the support of many concomitant observations. The mass flow theory is mainly based on the extreme concentration gradient between the source and the destination-called Sink. During translocation organic solutes enter from the mesophyll cells into sieve tubes; with the entry of food material high DPD gradient develops. As a result of this, water from the surrounding xylem cells enters into sieve tube cells, thus causing high degree of hydrostatic pressure. This pressure is the motive force for the movement of food materials enmass towards the least concentration/pressure region. This mass or pressure flow envisages unidirectional movement of materials.

This theory is supported by experimental evidences. If two membranous bags called A and B filled with concentrated and dilute sugar solutions respectively, are connected by a narrow glass tube and immersed in water, the solvent water enters into bag A due to a steep DPD gradient. As the water enters into the bag A, a very high hydrostatic pressure develops, with the result, sugar solution starts moving enmass into the glass tube towards bag B.

This experiment is further supported by the observation of high hydrostatic pressure in phloem elements. However, the rate of movement of solutes by pressure flow has been found to be extremely slow when compared to the actual rate of translocation observed in vivo. Added to this the unidirectional flow as envisaged by Mass flow contradicts the suspected bi-directional flow found in plants. The pressure of sieve plates offers greater resistance for the mass flow at the observed rate. At the top of it, mass flow never required ATP into consideration at all.
Basing on the above critical observations, it can be concluded that the over all mechanism takes place in two distinct but successive stages. The first is vein loading and the second is translocation through sieve elements.
Vein loading
Though carbohydrates are synthesized in chloroplasts during day time, very little of it is translocated and the rest is converted and stored in the form of starch. With the onset of night (darkness) starch hydrolysis gets activated (hydrolysis of starch into monosaccharides like glucose phosphate units which moves out into cytoplasm).
Light
Starch-> light+Pi----à Glucose-phosphates
Half the amount of the glucose-phosphate will be converted into Fructose 6-phosphate and the other half into UDPG.
Glucose 6 PO4 à Fructose 6-Phosphate
Glucose 6-PO4 + UTP à UDPG.
Fructose-6-phosphate and UDPG, in the presence of sucrase synthetase, combine to form sucrose-p. The probable role of Boron is believed to be the activation of sucrase synthesis.
Fructose-6P + UDPG à Sucrose-P
Sucrose phosphates are picked up by specific carriers. The same is actively transported across the cells and unloaded into sieve tubes found in the veins. Some of the cells, found around phloem elements act like transfer cells in transporting the sucrose into sieve tubes. This process is called vein loading. Inhibition of energy supply prevents vein loading which indicates that this process is an active process.
Development of Hydrostatic pressure or Mass Pressure
The loading of sucrose and other active substances into sieve tubes results in the high osmotic pressure. As a result of steep DPD gradient created between the water containing xylem cells and sucrose laden sieve tubes. Automatically, water diffuses into sieve cells in greater amount of bring about equilibrium, but it cannot do so because of the continuous input of the sucrose. This rush of water into translocation tubes causes enormous hydrostatic pressure, which pushes or forces the entry of solutes into other sieve tubes where pressure is very low. This acts as the beginning of a long journey of solute transport.
Further transport
Once the mass pressure builds up, the solutes have no other avenue but they have to move into other cells with low osmotic potential. But the sieve plates act as obstructive structures for the enmass movements. The solutes have to cross innumerable sieve plates found in the pathway from the source to sink. This obstruction has been found to be overcome by the P-proteins that are found traversing sieve plates. Being micro-tubular in nature, the P-proteins act like channels for the smooth flow of their activation by ATP molecules. Recent studies, under high voltage electron microscope show that the microtrabeculae play an important role in transportation of solutes.
This comprehensive explanation satisfied all the observed facts and about solute translocation like, energy requirement, involvement of P-proteins and hydrostatic pressure, as the motive force for the higher rate of translocation.