PLANT ANATOMY
The science of the structure of the organized plant body, learned by dissection, is called Plant Anatomy (Anatomy-dissection). In general, Plant Anatomy refers to study of internal morphology, pertaining to different tissues. The subject of this chapter is structure of Angiosperms, with emphasis on primary tissues.
TISSUE SYSTEM
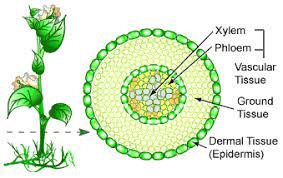
Plant body in Angiosperms is differentiated into root, stem, leaf and flower. All these parts are made up of different types of tissues containing different cell types. A tissue is a mass of similar or dissimilar cells performing a common function.
The body of a vascular plant is composed of Dermal tissue, Ground tissue and Vascular tissue.
Dermal Tissue (Skin):
Dermal Tissue is protective in function. Basing on its origin, it is classified into two types – Epidermis and Periderm.
Epidermis: This is the primary surface tissue of the entire plant body. Epidermal cells are compactly and continuously arranged on the surface of stem, root and leaves and many other structural organs; the continuity is lost by the presence of Stomatal pores or breaks in the tissue. Covering the aerial epidermis, cutin (fatty substance) is present as an impregnation on cell wall. The cuticle can be separated from epidermis. The epidermis may produce unicellular or multicellular hairy outgrowths and other appendages. Epidermis provides mechanical protection, allows gaseous exchange through stomata, restricts transpiration with cuticle, and is also involved in storage, photosynthesis, secretion, absorption and perception to stimuli. Stomata are Pores, each guarded by two guard cells, which control the size of the pore. Cells surrounding guard cells, but differing from other epidermal cells, are called subsidiary cells. Guard cells are kidney shaped; their cell walls are thick on the inner surfaces. Guard cells contain many chloroplasts.
Periderm: This is formed during secondary growth replacing primary epidermis. It is many layered.
Ground Tissue
This is inner to dermal tissue and is composed of simple tissues like parenchyma.
https://www.toppr.com/guides/biology/anatomy-of-flowering-plants/tissue-systems/
Vascular tissue
Vascular tissue consists of conducting elements – xylem and phloem. Vascular tissue may be scattered in ground tissue or regularly arranged forming a ring. In the latter arrangement, ground tissue is differentiated into cortex (outer to vascular tissue) and pith (inner to vascular tissue). The ground tissue of leaves is called mesophyll, bound by upper and lower epidermis.
TISSUE TYPES
In broad sense, tissues are classified as – meristematic and permanent tissues.
Meristem (Meristos – divisible).
Initially all embryonic cells of an embryo have the capacity to divide and multiply but as the embryo develops into a plant body, this capacity for division is restricted to certain parts of the plant body called meristems which are active throughout the life of the plant body (unlike that of an animal body). When meristematic cells divide, a group of the daughter cells remain meristematic; the other daughter cells called derivatives differentiate into various tissue elements. Before the occurrence of any cell division, usually cells become enlarged accompanied with addition of protoplasmic and cell wall material.
![]()
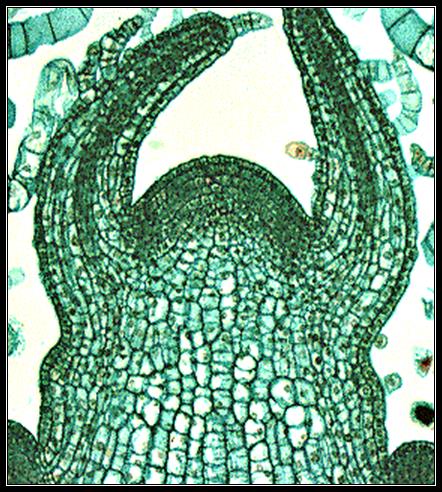
Meristematic cells are isodiametric, compactly arranged with dense cytoplasm, large nucleus, and small vacuoles or without vacuoles. Cell walls are thin.
Meristems which occur at the apices of stem, root and other branches are called apical meristems, which bring about primary growth of the plants, hence also called as primary meristems. In many plants in addition to apical meristems, lateral meristems like vascular cambium, cork cambium, intercalary meristems are found. Lateral meristems are arranged parallel to the sides of organs in which they occur.
Intercalary Meristem: This is also a primary meristem, found in permanent tissues; in the bases of internodes and leaf sheaths of grasses and many others. Sometimes, as in gynophore of groundnut, xylem may be present in intercalary meristem. Wherever stem is jointed, elongation of internodes is due to intercalary meristem. Example: Bamboos. Even prolonged growth of leaves, flowers and fruits may be regarded as an intercalary growth.
Vascular cambium and cork cambium: These are referred to as secondary meristems because they produce secondary tissues, and increase the thickness of the organs and the plant body. This process is called secondary growth, seen in dicotyledons and gymnosperms.
Permanent or Mature Tissues
Change in their structure, metabolism and chemistry and acquire specialized characters by their various modes of differentiation. Not all the cells totally differ from the meristems. Some cells retain the power of cell division and others cannot divide. In a strict sense only cells which have lost the power for division must be regarded as permanent tissues, but in a broad sense, cells derived from meristem that have acquired a special functions like photosynthesis, secretion, storage are treated as part of matured tissue. There are different types of mature tissues. Example: Parenchyma, Collenchyma, Sclerenchyma, Xylem and Phloem.
Types of Mature Tissues
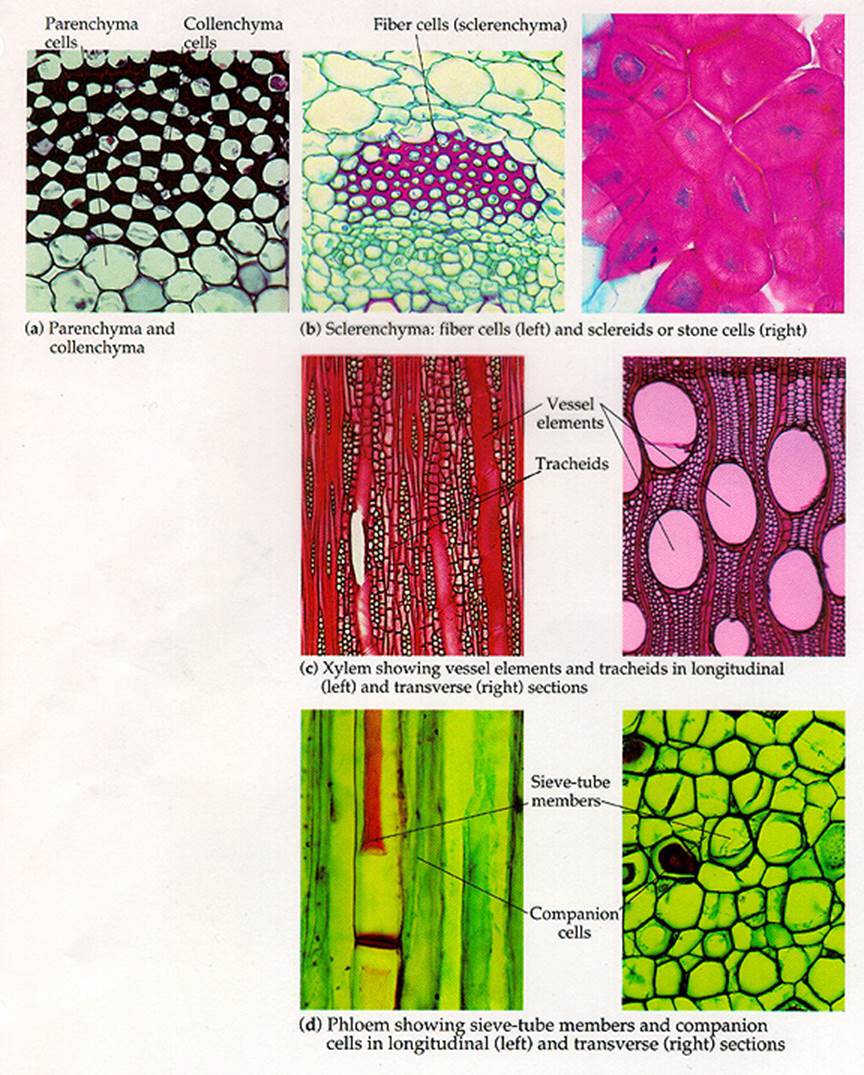
Parenchyma (Para-besides, enchyma-In poured)
Parenchyma is the fundamental tissue of the plant body. It is found in every part of the plant body like pith and cortex of stem and root, mesophyll of leaves, flesh of fruits, floral parts and even in xylem and Phloem. Cells have thin primary walls and polyhedral shapes. The average number of faces of a polyhedron is 14 and is called as "tetra decahedron". Cells are compactly arranged or more commonly spaciously arranged with intercellular spaces as in cortex and pith. Cells possess dense cytoplasm and are active metabolites.




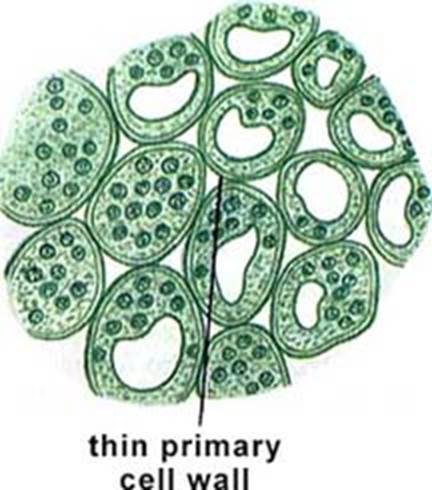
Generally, cells of parenchyma are involved in storage of starch, sucrose, protein, water, phenol derivatives and many mineral substances. Other metabolisms like respiration, protein synthesis etc., are active in the tissue. Parenchymatous cells may also perform specialized functions and are structurally modified. The following are the different types of parenchyma.
Aerenchyma: Intercellular spaces filled with air, are large in size and many in number. Cells occupy a smaller area. Though the cells are smaller, they provide the required strength to the aquatic plants. In these plants air spaces are common, helping in aeration and buoyancy. Air spaces are also seen in roots of grasses, petioles of canna, aroids etc.
Chlorenchyma: Cells of photosynthetic parenchyma contain numerous chloroplasts. These cells are commonly seen in leaves, some times in young shoots. Cells of Chlorenchyma are of two types –
1) Palisade cells that are elongated and compactly arranged.
2) Spongy cells that are spaciously arranged and irregularly shaped.
Prosenchyma (Pros-to): Cells are elongated, have thick walls and serve as a supporting tissue. Example: Endosperm cells of seeds.
Armed Parenchyma: Cells are stellate in shape with inward cell wall projections. Example: Pinus leaf Mesophyll.
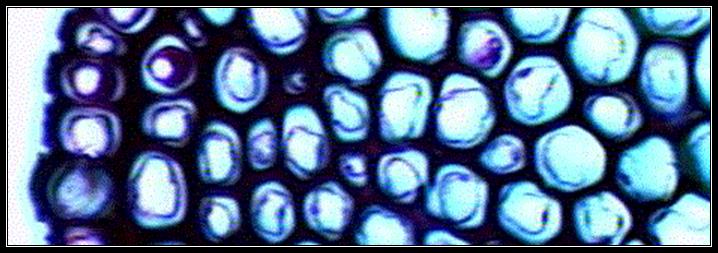
Collenchyma (Kolla – Glue):
Collenchyma is a simple, living tissue. Cell walls are thickened due to deposition of pectin. Collenchyma is the primary supporting tissue in stems, leaves and floral parts of dicots, where as in stems and leaves of monocots collenchyma is usually absent, (instead, sclerenchyma is present in monocots).
· 
![Text Box: [Grab your reader’s attention with a great quote from the document or use this space to emphasize a key point. To place this text box anywhere on the page, just drag it.]](3_PLANT_ANATOMY_files/image013.gif)

Collenchyma is usually hypodermal in position. Cells are more elongated and narrower than parenchyma. Like Parenchyma, collenchyma may also contain chloroplasts or may regain the thickening. Intercellular spaces may or may not be present. Three forms of collecnchyma are recognized based on the types of thickenings –
1. Thickening is on the tangential wall – lamellar collenchyma.
2. Deposition of pectin is in the corners where several cells meet-angular collenchyma.
3. Thickenings are around the intercellular spaces – lacunar collenchyma.
Being plastic, collenchyma differs from elastic fibers of sclerenchyma.
Sclerenchyma (Scleros-hard)
Cells of sclerenchyma are thick walled and are usually lignified. The thickness is due to formation of secondary wall. At least initial the secondary wall is free from primary wall. At maturity, usually the cells are devoid of protoplast. The cell wall encloses a cavity lumen. On the cell wall, pits are usually present (simple type). Commonly sclerenchyma cells are classified into fibers and sclereids. The primary function is mechanical. (Pits-thin areas without secondary wall material deposition).
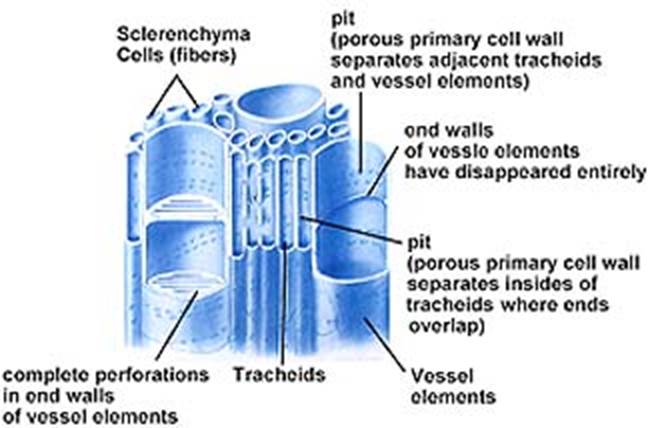
Fibers: These are usually long, spindle shaped structures, with tapering or blunt ends. Longest fiber is seen in Boehmeria nivea (55 cms). They are arranged in groups. Secondary thickening may account for 90% of the area of the cell; the lumen is narrow. Fibers are grouped into xylary fibers and extraxylary fibers. Xylary fibers, also called wood fibers are parts of xylem and are longest among xylem elements. Extraxylary fibers are classified as – Bast or phloem fibers, cortical fibers and perivascular fibers (peripheral to the vascular bundles). These are lignified or non lignified.
On account of their elasticity, fibers enable the plant body to withstand various strains. Commercial fibers like jute, flax, ramie are extraxylary fibers.
Sclereids: These are shorter than fibers. Sclereids occur singly or in groups. Sclereids are commonly found in fruit wall, seed coat, epidermal scales, and occasionally found in cortex, pith, mesophyll and petiole of submerged aquatics.
There are many types of sclereids –
1. Asterosclereids are star shaped
2. Macrosclereids are similar to palisade cells
3. Osteosclereids are bone like that are enlarged at their ends
4. Brachysclereids are isodiametric like parenchyma
Vascular Tissue
This tissue is also called complex tissue. It is heterogeneous in nature with different types of cells. The chief elements are xylem, phloem and pericycle.
Xylem (Wood):
Xylem is the water conducting tissue. It consists of living cells like parenchyma and dead cells like tracheary elements. Fibers are also present. Xylem elements differentiating from an apical meristem constitute primary xylem where as secondary xylem is differentiated from vascular cambium. Xylem is the most preserved tissue in fossils, due to the development of lignified secondary walls.
Tracheary elements (trachea – rough): These are of two types – 1). Trachea or vessels 2) Tracheids, Vessels are limited in their growth, are joined end to end to form continuous tubular structures with perforations in their cross walls. Water and minerals are efficiently conducted through these perforations. Vessels are present in most of the Angiosperms and also in certain lower plants like Gnetum, Marsilea and Selaginella.
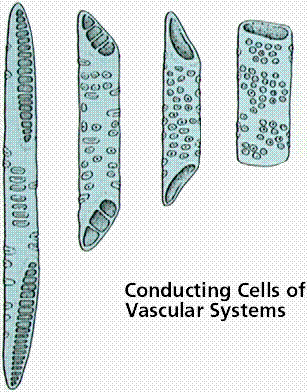
TracheidsTracheids
are generally elongated and non-perforated. They have only pit pairs, at the
regions of union with other tracheids. Conduction of water and minerals is not
as efficient as in vessels. Tracheids are found in Pteridophytes, most of the
Gymnosperms and a few Angiosperms.
Secondary thickenings in tracheary elements are deposited in various forms-1) as rings, called annular. 2) as continuous helices, called helical or spiral, 3) as net work called reticulate, 4) as transverse nets called scalariform, 5) as extensive thickenings except in the region of pits, called pitted.
Primary Xylem: This is differentiated into Protoxylem and Metaxylem, denoting the xylem that appears first. In stems, protoxylem is found nearest to central axis (endarch xylem) and in roots away from centre (exarch xylem). The protoxylem elements have annular, spiral and sometimes reticulate secondary thickenings. Fibers are absent in protoxylem. Metaxylem appears after protoxylem. Metaxylem has only pitted secondary walls. Metaxylem is more complex than protoxylem and its elements are wider.
Secondary Xylem: The vascular cambium (intrafascicular cambium interfascicular cambium) producing secondary xylem consists of fusiform and ray initials. Here also the secondary xylem is also of two-system-1) Axial or vertical system with living and nonliving cells and 2) ray or horizontal system with living cells. When compared with primary xylem, secondary xylem is more complex and shows orderly development. Pitted and scalariform secondary thickenings are developed.
Phloem (Phloos Bark)
Phloem is the food
conducting tissue. It is composed of sieve elements, parenchyma cells, fibers
and sclereids. Phloem differentiateddifferentiated
from procambium is called primary phloem; secondary phloem is initiated from
vascular cambium.
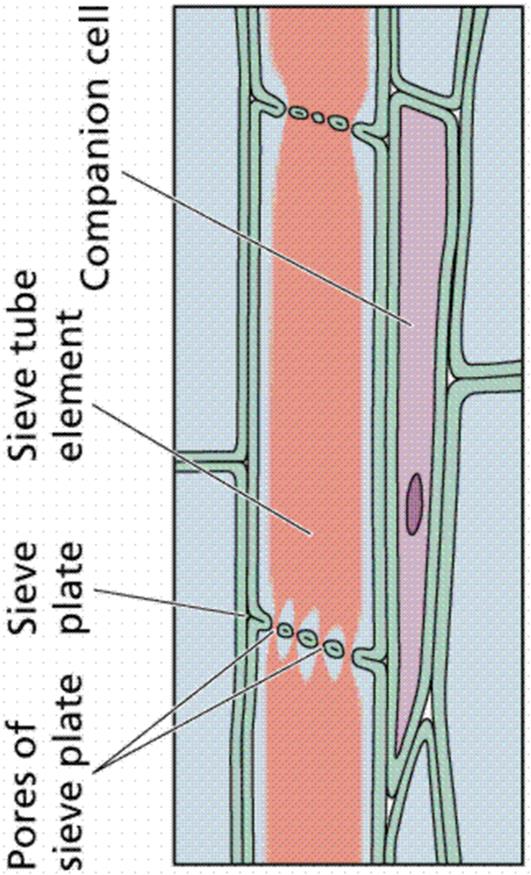
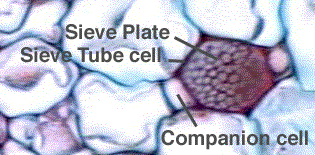
When xylem and phloem elements are found in the same line the arrangement is called conjoint; if phloem is only external to xylem it is called collateral as in many stems; if phloem is both external and internal to xylem it is called bicollateral as in leaves and a few stems; if xylem completely surrounds phloem it is called amphivasal; if phloem surrounds xylem it is called amphicribal; if xylem and phloem alternate with each other as in roots, it is called radial.
Sieve Elements: These are classified into sieve cells and sieve tube members. Sieve cells are commonly long, slender, and taper at both ends. These cells overlapping, many sieve areas are seen. Sieve areas have cluster of pores through which adjacent sieve cells are interconnected by protoplasmic strands. Surrounding the pores, carbohydrate Callose material is found in a sectional view. Sieve cells are found in Pteridophytes and Gymnosperms. Sieve tube members are tubular, placed in long series and have specialized sieve areas called sieve plates occurring at end walls. These sieve plates are often inclined. In addition to sieve plates, there may be also sieve areas on lateral walls of sieve tube members. In sieve plate, sieve areas, having large pores are seen. Initially sieve tube members contain uninucleated protoplast. Gradually the nucleus and its associated endoplasmic reticulum become disorganized. Mitochondria may lose their inner membranes. Cytoplasm is usually parietal. In the centre of the cell a mixture of vascular sap and disorganized cytoplasmic matter is seen. Tonoplast is not found surrounding the vacuole. Studies sieve contents show that they are under positive pressure of 30 atm and velocity of conduction is 100 cm/ hour. The transported material is usually sucrose which is more than 20% in concentration.
Sieve tube member is associated with one or more specialized parenchyma cells called companion cells are seen only in Angiosperms. Both sieve tube member and companion cell are derived from the same meristematic cell. Often, companion cells of sieve tube members form longitudinal series. Companion cell is nucleated. Probably these cells provide energy for food conduction.
Primary Phloem: This is classified into Protophloem and Metaphloem. In protophloem, sieve tube members are without companion cells. Sieve tubes function for a brief period and soon they get crushed by the surrounding pressure. The crushed cells may disappear. Metaphloem tissue survives for a longer period. Its elements are longer and wider. Usually fibers are absent in dicotyledons, where as in monocotyledons and herbaceous dicots, parenchyma cells are absent.
Secondary phloem: Similar to secondary xylem, there are two systems of arrangements in secondary phloem – 1) Axial system producing sieve elements, phloem parenchyma and phloem fibers. 2) Transverse system producing ray parenchyma cells.
Pericycle (A Circle All Around)
Pericycle is the region, consisting of one – few layers of cells, found external to central cylinder (Stele). Example: Roots and Dicot stems.
Pericycle may be composed of parenchyma cells or sclerenchyma cells or both. Lateral branches and phellogen may arise from pericycle.
Summary
Tissue: a group of cells performing a particular function is called tissue. Basing on the types of cell structure and function, tissues are classified into Meristematic, Simple and Complex tissues.
Meristematic Tissues: These are actively dividing cells, which are isodiametric in shape, rich in cytoplasm, with small vacuoles or no vacuoles and cells are actively metabolic. Depending upon the position they are classified into apical meristem (at the tips of the stem and roots), lateral meristem (vascular and cork cambium), intercalary meristem (at the basal portion of internodes in Graminae members). Apical meristem is responsible for the primary growth, ex., growth of the plant in length; lateral meristem is responsible for secondary growth and intercalary meristem is responsible for the elongation of the internodes in Bamboo plants etc.
Simple Tissues: Tissues contain cells of similar structure, function and have common origin.
Method of study: Stem cuttings are put in a macerating fluid and allowed for some time; meanwhile some think T.S. are taken and stained with saffranin and mounted on a slide. The treated stem pieces are taken on the slide, washed, teased and mounted on the slide for microscopic observation.
Parenchyma: Important living tissue found abundantly in cortex, pith of the stem, root and also in the mesophyll tissue of leaves. Depending upon the structure, content and function they are differently named like storage tissue, chlorenchyma, spongy parenchyma, ground tissue, aerenchyma etc. Cells are thin walled, rich in cytoplasm, with small or a large central vacuole, actively metabolic and loosely arranged with small or large intercellular spaces.
Collenchyma: Important Living mechanical tissue, found mostly just below the epidermis of stem, similar to that of parenchyma but with difference in the cell thickenings. Here cells are larger, compactly arranged and cell wall at the angles of the meeting of cells is characteristically thickened by pectin deposition.
Sclerenchymatous Fibers: These are true mechanical tissues either dead or living. Two types of cells are found a) Sclerenchyma fibers b) Sclereids.
Sclerenchymatous Fibers: They are dead cells, long with tapering ends, with uniform thickening, leaving a narrow lumen. They are found below the epidermis, cortex, around vascular bundles and so on.
Sclereids: They are found in fruit or seed walls and also in the flesh of apples and pineapple. The cells may be living or dead, but short and variously shaped; cell wall is heavily thickened with many pit lines unevenly distributed.
Complex Tissues: The cells that are grouped here are of various types in their structure, shape and function, and have different origin, but together they perform a common function Example: vascular tissues, secretory tissues etc.
Vascular Tissues: 1) XYLEM: It is a water conducting tissue, made up of tracheids, trachea, xylem fibers and xylem parenchyma of which the first three are dead cells and the last one is living.
TRACHEIDS: have tapering ends with secondary thickenings of annular, spiral, scalariform and reticulate types and cell walls are pitted.
TRACHEA: are vessels with the tapering ends and their transverse walls are disintegrated, so that cells are placed end to end to form a kind of continuous channels for conducting water and minerals.
XYLEM FIBRES: are similar to Sclerenchymatous fibers. All the above three tissues are dead and by having thick secondary walls they act as mechanical tissues.
XYLEM PARENCHYMA: even though its cell walls are thick and pitted it is living and perform some visit functions probably active transportation etc.
3) PHLOEM: This tissue is solely responsible for the transportation of the food material from the source i.e., leaves and storage organs to the regions of need. It consists of sieve tubes, companion cells, phloem parenchyma (all living) and phloem fiber (dead cells). SIEVE TUBE CELLS : are long large cells with end walls perforated, but cushioned with callus material and the end plates are called sieve plates, through which large number of cytoplasmic strands pass through. The cell has a large central vacuole with thin peripheral protoplasm. The most remarkable feature is the complete absence of nucleus. Most of the food material is conducted through this sieve tube in the form of sucrose.
4) COMPANION CELLS: are small cells, living, found and pressed to sieve tubes. Cells are rich in cytoplasm and actively metabolic. The function is probably providing energy for transportation of food materials by active process. PHLOEM PARENCHYMA: is similar to parenchyma, and found around sieve tubes and companion cells PHLOEM FIBRE OR BAST FIBRES: are mostly dead cells with considerable and varied type of secondary thickenings.
PRIMARY GROWTH
The primary growth of a plant body is brought about by the activity of primary meristems, particularly by apical meristem. Details about primary growth occurring in stems, leaves and roots, may be studied by observing a transverse structure is differentiated into outer epidermis, the inner ground tissue and vascular tissue.
Dicotyledonous stem: Example: Sunflower (Helianthus annus).
The transverse section is almost circular in outline.
Epidermis: This is the outermost layer, uniseriate, covered with thin cuticle, and bears a few stomata and many multicellular trichomes.
Ground Tissue: This fundamental tissue is differentiated into outer cortex and central pith.
Cortex: This is many layered and differentiated into hypodermis and inner cortex. Hypodermis is found below the epidermis, and is constituted with angular collenchyma which is 3-5 layered.

The inner part of cortex is constituted with chlorenchyma and parenchyma; large numbers of intercellular spaces are present between these cells. The inner most cortical layer is wavy in outline, rich in starch grains, with barrel shaped cells and is called endodermoid layer. (Endodermis is generally absent in stem; endodermoid layer differs from endodermis in lacking casparian thickenings).

Pericycle: This is the outermost region of vascular tissue. This consists of alternately arranged patches of sclerenchyma and parenchyma. The sclerenchyma is also known as hard bast fibers. Since it is found external to vascular bundle, the patch of sclerenchyma is also called bundle cap.
Vascular tissue: This consists of many vascular bundles arranged regularly in the form of broken ring (Eustele). In each vascular bundle the xylem and phloem elements are in the same line; the presence of one to few stripped intrafascicular cambium is found in between xylem and phloem; the cambial cells are thin walled and are almost rectangular phloem is only external to xylem: the protoxylem is nearest to centre. The vascular bundles are conjoint, collateral, endarch, open and wedge shaped.
Pith or Medulla: This is inner to vascular tissue and occupies the central region. It is composed of cells of parenchyma, with intercellular spaces. Extensions of these cells into the areas in between vascular bundles are called medullary rays.
Monocotyledonous stem: Example: Zea mays (Maize). The outline is circular.

Epidermis: This is the uniseriate, outermost layer and is covered with thick cuticle. Trichomes are absent.
Ground tissue: The ground tissue is not differentiated; into cortex and pith, since there is scattered arrangement of vascular bundles. Even endodermoid layer and pericycle are also not differentiated. Hypodermis constitutes the exterior ground tissue, and is composed of 2 – 3 layers of sclerenchyma. The major part of the ground tissue is parenchymatous.
Vascular tissues: This consists of numerous, oval shaped vascular bundles scattered throughout the ground tissue, hence the arrangement of vascular tissue is called atactostele. Peripheral vascular bundles are small-sized but many in numbers. The central vascular bundles are large sized but a few in numbers in all the vascular bundles the xylem and phloem are in the same line; cambium is absent; phloem is only external to xylem; protoxylem faces the centre. Hence the vascular bundles are described as conjoint, collateral, endarch and closed. Each vascular bundle is surrounded by a bundle sheath of sclerenchyma, which is prominent at the base and tip of vascular bundle. Xylem comprises usually 2 oppositely arranged Metaxylem elements (pitted, facing periphery) and a few protoxylem elements (facing the centre of stem) in a linear row. If an imaginary line is drawn connecting the two Metaxylem elements with that of protoxylem, xylem appears ‘Y’ shaped. Usually the lower protoxylem elements break down and form a lysogenous cavity. Above the metaphloem, a small band of obliterated protophloem is seen. Metaphloem consists of sieve tubes and companion cells but not phloem parenchyma.
Comparison between the stems of Dicot and Monocot:
|
|
Dicot stem Ex: Sunflower |
Monocot stem Ex : Maize |
|
1. Epidermal trichomes |
Present multicellular |
Absent |
|
2. Hypodermis |
Collenchymatous |
Sclerenchymatous |
|
3. Ground tissue |
Differentiated into cortex and Pith |
Undifferentiated |
|
4. Endodermoid layer and Peri-cycle |
Present |
Absent |
|
5. Stele |
Eustele |
Atactostele |
|
6. Vascular bundles |
Many open, wedge- shaped |
Numerous, closed, oval-shaped |
|
7. Lysigenous cavity |
Absent |
Present |
|
8. Bundle cap |
Present |
Absent |
|
9. Bundle sheath |
Absent |
Present |
|
10. Secondary Growth |
Occurs later |
Does not occur |
Dicotyledonous root: Example: Lablab purpureus (Beans)

Epiblema: The external protective layer is called Epiblema or Piliferous layer (the term epidermis is generally not applied to roots). It is uniseriate. Cells
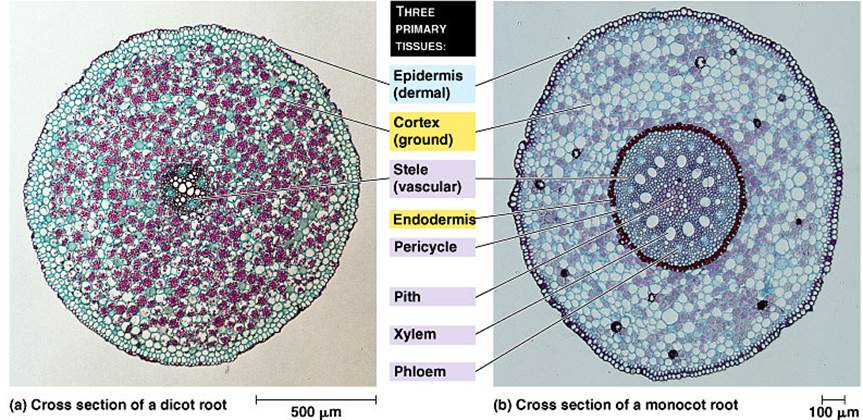
are thin walled. Some of these cells extend into unicellular root hairs.
Cortex: The region inner to epiblema is called cortex and it is homogenous. The cortex is composed of thin walled, spaciously arranged parenchyma cells that constitute many layers. With increase in age, the exterior cortical region becomes suberized and dead. This exterior cortex is known as exodermis. The innermost cortical layer is called endodermis. Its cells are barrel-shaped and are closely packed. Many cells get suberized on their radial and inner walls. These thickenings are called casparian stripes. Such endodermal cells may become totally suberized on all wall surfaces and are called stone cells. These cells do not allow conduction. The non-suberized endodermal cells are called passage cells. Inner to endodermis, one or two layers of parenchyma are present, constituting pericycle.
Stele: Xylem and phloem patches are in equal number and alternate with each other. Xylem or phloem patches are usually four in number. In Xylem, the protoxylem elements face pericycle; between xylem and phloem elements, parenchyma cells are present constituting conjunctive tissue, which later becomes a secondary meristem. Hence vascular bundles are described as radial, exarch and open.
Pith is very small, with closely packed parenchyma cells.
Monocotyledonous root: Example: Canna
Outline almost circular.
The nature of epiblema, exodermis, cortex, endodermis and pericycle is similar to that of Dicot root. However in cortex, large air spaces of schizogenous origin may be seen. The nature and arrangement of vascular bundles are also similar to that of Dicot root. But vascular bundles are usually more than six in number. The Parenchyma present in between xylem and phlegm is called conjunctive tissue, with thin cell walls. This tissue does not become meristematic and thus differs from Dicot root. Pith is relatively large and parenchymatous. The parenchyma of the
Pith and conjunctive tissue, become lignified and Sclerenchymatous. The vascular bundles are described as polyarch, radial, exarch and closed. An endogenous lateral branch may be seen developing from pericycle and passing through cortex.
Comparison between the roots of Dicot and Monocot:
|
|
Dicot Root Ex: Beans |
Monocot Root Ex : Canna |
|
1. Vascular bundles |
Diarch to hexarch, open |
Polyarch closed. |
|
2. Pith |
Small or absent |
Relatively large |
|
3. Cambium |
Conjunctive tissue becomes meristematic and results in secondary growth |
Conjunctive tissue does not become a meristem and complete absence of secondary growth. |
|
4. Pericycle |
It may give rise to cork cambium and lateral branches |
It may give rise to only lateral branches. |
Comparison between the structures of stem and root:
|
|
Stem |
Root |
|
1. Cuticle |
Present, thick / thin |
Usually absent |
|
2. Trichomes |
When present, is multi-cellular or unicellular on the Epidermis. |
Epiblema hairs are unicellular |
|
3. Stomata |
Present |
Absent |
|
4. Hypodermis |
Either collenchymatous or sclerenchymatous. |
Not distinct from inner cortex. |
|
5. Cortex |
When present, is heterogeneous, with collenchyma, parenchyma and sclerenchyma. |
Homogenous with cells of parenchyma. |
|
6. Endodermis |
Usually absent; endodermoid layer may be present |
Generally present |
|
7. Pericycle |
Two to few layered/absent |
One layered |
|
8. Vascular bundles |
Many, conjoint, collateral or bicollateral, regularly arranged forming a broken ring or scattered in ground tissue, endarch. |
2-12, radial, exarch, form a broken ring. |
|
9. Pith |
Well marked and large or not differentiated |
Relatively small or absent. |
|
10. Branching |
Exogenous, originating from cortex |
Endogenous, arising form pericycle. |
Dicotyledonous leaf: Example: Mangifera (Mango)
The leaf is horizontal and dorsiventrally differentiated (upper = ventral, lower = dorsal).

Epidermis: The two surfaces are bounded by their respective epidermal layers. The epidermal cells are barrel-shaped, compactly arranged; upper epidermis is covered with thick cuticle and lacks stomata; lower epidermis is light green, covered with thin cuticle and is interrupted by stomata.
Mesophyll: This is the ground tissue. It is differentiated into upper palisade and lower spongy parenchyma.
Palisade parenchyma is found immediately below the upper epidermis, 2 to 3 layered, with compactly arranged tubular cells, rich in parietal chloroplasts.
Spongy parenchyma is found above the lower epidermis; these cells are varied in shape and sizes, very loosely arranged enclosing air spaces some of which open into stomata. Chloroplasts are parietal in the parenchyma cells.
Vascular tissue: Vascular bundles vary in size; each bundle is conjoint, collateral and closed. The vascular bundle is covered by a bundle sheath of parenchyma cells. An extension of bundle sheath is seen above and below the vascular bundle, reaching the epidermal layers. Phloem is towards lower epidermis; xylem is towards upper epidermis, with metaxylem facing phloem. Fibers are absent in both xylem and phloem. The xylem and phloem elements are conspicuous only in large vascular bundles.
Monocotyledonous leaf Example: Maize.
The leaf is Isobilateral in nature and is vertically oriented.

Epidermis: This is uniseriate, with barrel-shaped, compactly arranged cells and is covered with thick cuticle.
Stomata are found on both upper and lower epidermal layers hence it is called amphistomatic (more on the lower epidermis). Though the leaf is referred to as isobilateral, it is only in upper epidermis, a few large, empty and colorless bulliform or motor cells are present. During dry weather, these motor cells help the leaf to role over, due to the changes in turgidity. This rolling of leaf reduces the rate of stomatal transpiration.
Mesophyll: There is no differentiation of mesophyll into spongy and palisade parenchyma. All the cells of chlorenchyma are alike, isodiametric, almost compactly arranged with numerous parietal chloroplasts.
Vascular tissue: The vascular bundles are numerous, arranged in parallel series (venation is palmate-parallel), conjoint, collateral and closed. Phloem is towards lower epidermis. Each vascular bundle is surrounded by chlorenchymatous bundle; this sheath also serves for temporary storage of starch. A few vascular bundles are larger in size, with more amount of xylem and phloem and with large bundle sheath cells. A patch of sclerenchyma is present above and below the large sized vascular bundles.
Comparison between the leaf structures of Dicot and Monocot.
|
|
Dicot Root Ex: Mango |
Monocot Root Ex : Maize |
|
1. Orientation of leaf |
Horizontal, dorsiventrally differentiated |
Vertical, isobilateral |
|
2. Stomata |
Usually on lower epidermis |
On both epidermal layers |
|
3. Cuticle |
Thick on upper epidermis |
Thick on both epidermal layers |
|
4. Motor cells |
Absent |
Present in upper epidermis |
|
5. Mesophyll |
Differentiated into upper palisade parenchyma and lower spongy parenchyma |
No differentiation of mesophyll |
|
6. Vascular bundles |
All of them are not seen in parallel series and are supported by bundle sheath extension |
Vascular bundles are in parallel series and are supported by sclerenchyma patches |
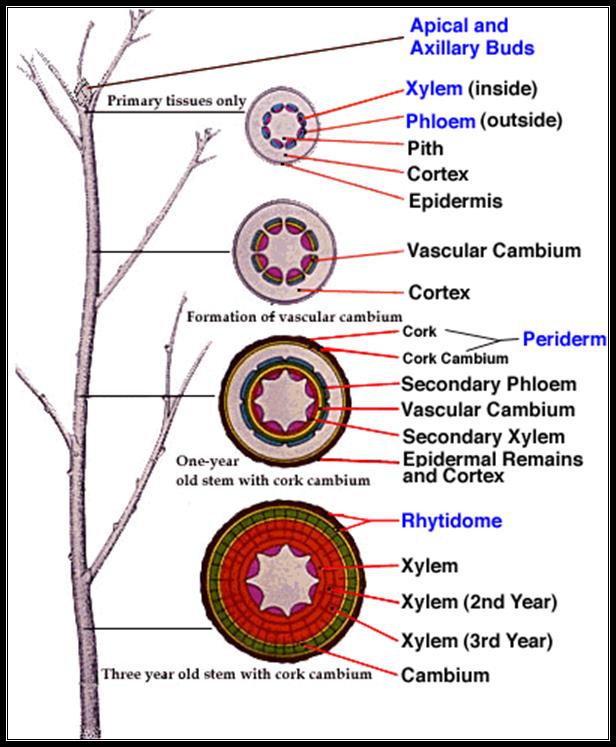
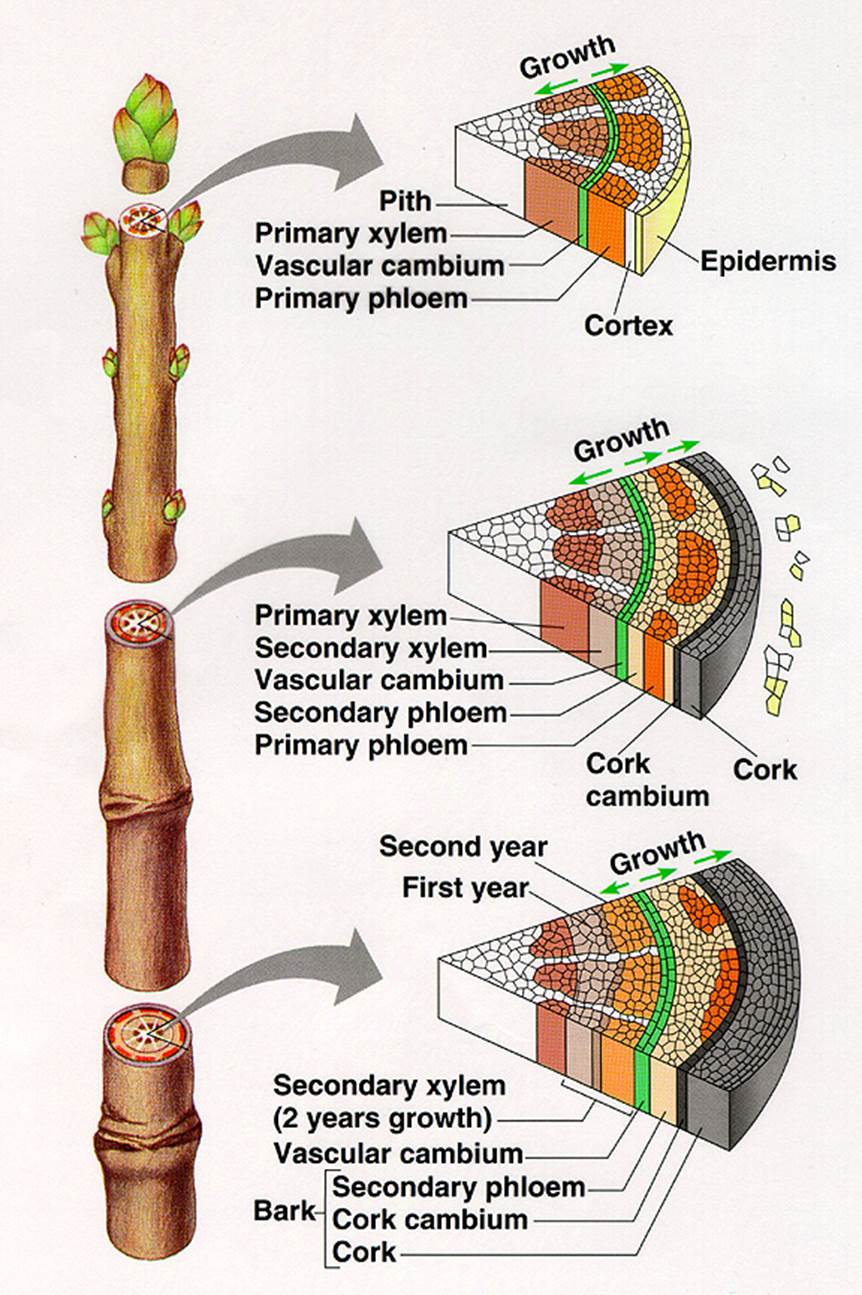
Secondary Growth in Dicot Stem
Increase in lateral thickening is called secondary thickening or secondary growth. This growth occurs in stem and root parts of shrubs and trees in Dicotyledons. This development takes place in stele (intrastelar secondary growth) and also in cortex (extra stellar secondary growth).
Intrastelar Secondary Growth: In each vascular bundle between xylem and phloem a strip o intra-fascicular (within vascular bundle) cambium is present. Between vascular bundles, medullary rays composed of parenchyma are found. A few parenchyma cells of these rays adjacent to intra-fascicular cambium dedifferentiate into meristematic cells and constitute inter-fascicular cambium (in between vascular bundle). Both inter-and intrafascicular cambiums join to form a complete ring of cambium called vascular cambium, which initiates intrastelar secondary growth.
The vascular cambial cells continuously divide and form numerous daughter cells. These derivatives produced inward towards pith mature into secondary xylem elements, while those produced outward toward cortex differentiate into secondary phloem elements. The amount of secondary xylem tissue produced is much larger than that of secondary phloem tissue. Due to the formation of secondary xylem and phloem, the primary xylem and phloem become separated. The primary phloem is pushed outward and is finally crushed due to internal pressure. The primary xylem is pushed towards the centre of the axis; however primary xylem elements are not crushed because the cell walls are lignified. Between bundles of secondary tissues parenchyma cells constituting vascular rays are found.
Extrastelar secondary growth: To keep pace with the expanding central stelar region and to maintain the area volume equilibrium between the stele and cortex, epidermis and pericycle, with parenchyma or collenchyma cells acquires secondary meristematic property by dedifferentiation and is called phellogen or cork cambium. This cambium is similar to vascular cambium in cutting off cells on its outer and inner faces. The outer derivatives gradually lose their protoplast and become dead; their cell walls become suberized. These cells constitute cork or phellem. These inner derivatives remain living and differentiate into parenchyma of collenchyma. This newly formed inner zone is called secondary cortex or phelloderm. The phelloderm, phellogen and phellem are the primary covering layer i.e., epidermis gets ruptured. The newly formed periderm becomes the protective layer.

Lenticels: the external phellem or cork is impermeable to conduction or exchange of gases; however it is not a continuous zone; a few gaps are seen in cork. In these gaps, a mass of parenchyma cells (complementary tissue) is seen. This structure is called lenticel. This helps in exchange of gases, between the atmosphere and inner cortical cells. Through lenticels also, transpiration occurs and is called lenticular transpiration (loss of water is insignificant).
Bark: In cortex of trees, generally many phellogens arise, each one producing outer phellem and inner phelloderm. If such a process takes place, whatever tissues lie outer to the inner most phellogen, become suberized and dead. This structure is called bark, which is of two types – 1) continuous bark 2) scaly bark or discontinuous bark. Bark acts as insulation. It is thicker in temperate tree forms.
Growth rings: In temperate woody plants and in a few tropical trees, the activity of vascular cambium is seasonal, but not uniform throughout the year. This is evident only in the region of secondary xylem. The cambium is less active in autumn, producing few small xylem elements that are having small lumen. This is called late wood or autumn wood. In spring the cambium is more active producing large number of large sized xylem elements that are less dense with large lumen. This is called spring wood or early wood. The difference in secondary xylem correlated to the cyclic seasonal changes; result in the formation of alternate rings of autumn wood and spring wood. Thus, in each year two zones of secondary xylem are formed and both can be easily demarcated. These two zones constitute an annual or growth ring. By counting the number of annular rings, it is possible to know the age of a plant. The availability of auxin may be regulating the activity cambium.
Wood: The elements of secondary xylem constitute wood. An active wood, containing water and stores food substances is called sap wood or alburnum. It is peripheral. It’s living parenchyma cells store and translocate food. Sap wood is soft and light in color.
 With advance in age, the central axis of
secondary xylem becomes inactive, losing water and stored food material. It
becomes infiltrated with various organic chemicals like oil and ergastic
substances (Passive byproducts of metabolism). This region becomes hard in
texture and dark in color and is called Heart wood or Duraman. This zone loses
the function of conduction of nutrients. Thus, an active wood in addition to
conduction of water also conducts nutrients.
With advance in age, the central axis of
secondary xylem becomes inactive, losing water and stored food material. It
becomes infiltrated with various organic chemicals like oil and ergastic
substances (Passive byproducts of metabolism). This region becomes hard in
texture and dark in color and is called Heart wood or Duraman. This zone loses
the function of conduction of nutrients. Thus, an active wood in addition to
conduction of water also conducts nutrients.