CELL DIVISION:
Cells of all organisms undergo cell division at one or the other stages of their development, even restoration of the lost cells, pre-existing cells divide and redivide and stop when the size of the tissue reaches the maximum or desired size. In many unicellular forms, cell division is an important mode of multiplication. But in multicellular organisms, cell division is absolutely required for growth. Reproductive elements like gametes are the other important products of cell division.
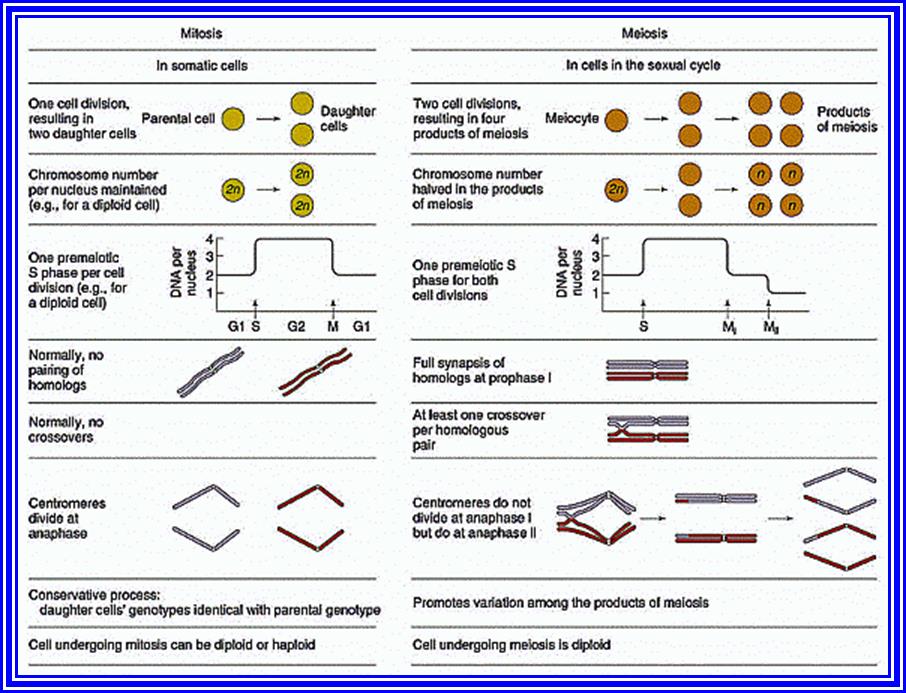
Types of Cell divisions: Organisms exhibit two types of cell divisions. This is based on the pattern of distribution of parental chromosomes to the daughter cells. They are Mitosis and Meiosis; however, in prokaryotic organisms like bacteria and blue green algae, where there is no organized nucleus, the cell division is equational and it is called Amitosis, for the mitotic apparatus and such complicated chromosomal movements are minimized or they reproduce by budding. Whatever may be the types; all cell divisions involve two important events like, nuclear division called Karyokinesis and cytoplasmic division called Cytokinesis.
Amitosis
Amitosis is also referred to as Binary fission or direct division. This type of division is employed by lower prokaryotes, not only for multiplication of cells but also for reproductivity. In this context, a bacterial cell has been taken as an example of describe the process.
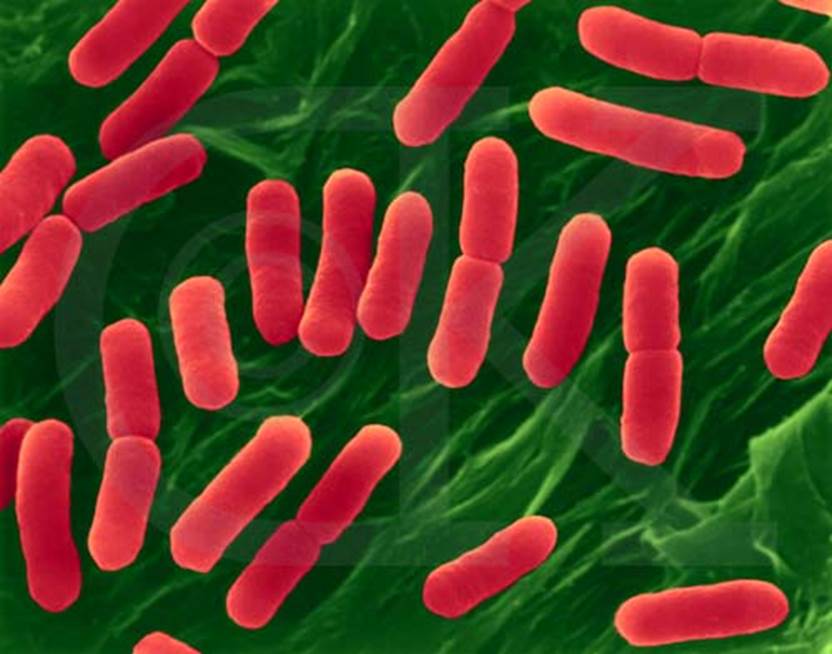
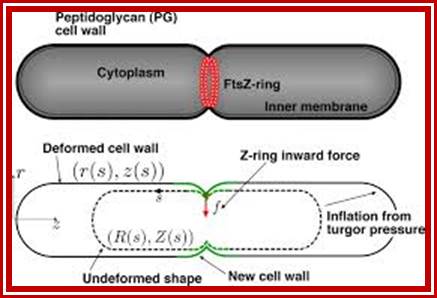
A mechanical model of bacterial cell division. The FtsZ-ring is positioned at the mid cell and generates a contractile force, f, in the inward radial direction (red arrow). The observed PG cell wall is inflated from the undeformed cell shape by internal turgor pressure, much like a balloon. The undeformed cell shape is defined by parametric profiles (R(s), Z(s)), which are the radial and longitudinal components of the material point coordinates; the deformed shape is similarly defined by profiles (r(s), z(s)); s is an arc-length measured from the mid cell. During division, deformed and undeformed shapes are changed by cell wall growth and remodeling (green regions indicate where most of the growth and remodeling occurs). Our model combines the mechanical properties of the cell wall with the growth and remodeling kinetics to compute the cell shape during division. https://www.pnas.org/
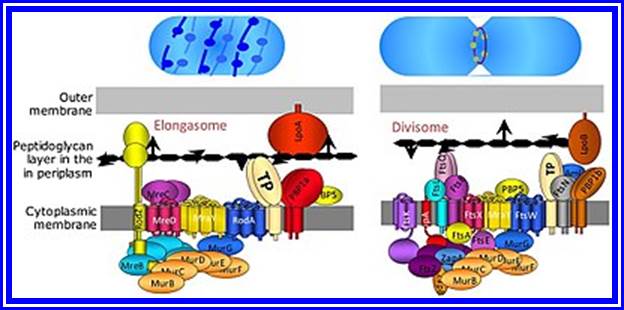
Divisome and elongasome complexes responsible for peptidoglycan synthesis during lateral cell-wall growth and division. Bacterial cell division happens through binary fission or sometimes through budding. The divisome is a protein complex in bacteria that is responsible for cell division, constriction of inner and outer membranes during division, and remodeling of the peptidoglycan cell wall at the division site. A tubulin-like protein, FtsZ plays a critical role in formation of a contractile ring for the cell division. https://en.wikipedia.org/
At the time of bacterial cell division, the circular DNA molecule that is attached to the mesosome membrane, starts replicating at the initiating point which is located very near to the mesosome.
The chemical components of mesosomes are responsible for initiating replication, which will be soon completed. Then the daughter molecules, again attach to the membrane, open out and segregate in a complex process. A little later, almost in the middle region of the cell, the plasma membrane produces an inward invagination all-round the cell and these progresses till the inwardly growing membranes fuse with one another in the center. This results in the division of cytoplasm into two compartments. Soon, between the newly formed plasma lemma, the cell wall materials such as peptidoglycans are secreted into the space found between them. Then the middle wall splits across in the middle and two daughter cells separate. At least 20 or more genes are involved in this process and it is highly regulated.
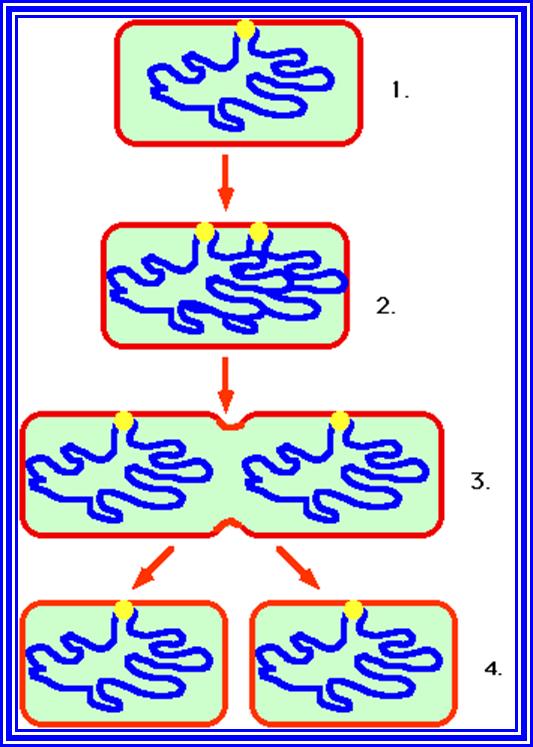
https://en.wikipedia.org/
The chemical components of mesosomes are responsible for initiating replication, which will be soon completed. Then the daughter molecules, again attach to the membrane, open out and segregate in a complex process. A little later, almost in the middle region of the cell, the plasma membrane produces an inward invagination all-round the cell and these progresses till the inwardly growing membranes fuse with one another in the center. This results in the division of cytoplasm into two compartments. Soon, between the newly formed plasma lemma, the cell wall materials such as peptidoglycans are secreted into the space found between them. Then the middle wall splits across in the middle and two daughter cells separate. At least 20 or more genes are involved in this process and it is highly regulated.
It is important to note that this type of cell division, though appears to be simple, it is complicated for the number of genes involved and the process involved.
MITOSIS
Multicellular organisms start their development as a unicellular 2n (diploid) cell or n (haploid) cell. Under favorable conditions unicellular organisms multiply and produce a huge population of cells. A fertilized egg in multicellular systems may develop into a giant plant. Some haploid organisms produce spores or gametes as a means of reproduction. All the above processes are achieved by mitotic divisions. This process involves equal distribution of genetic material.

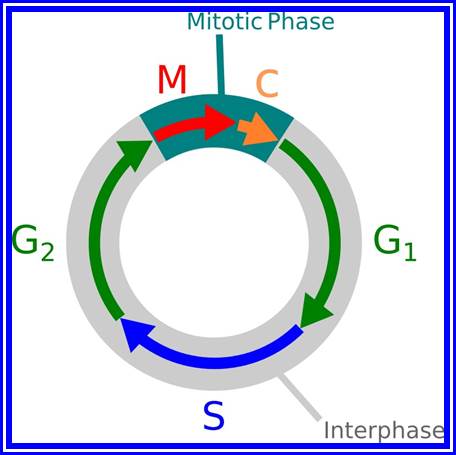
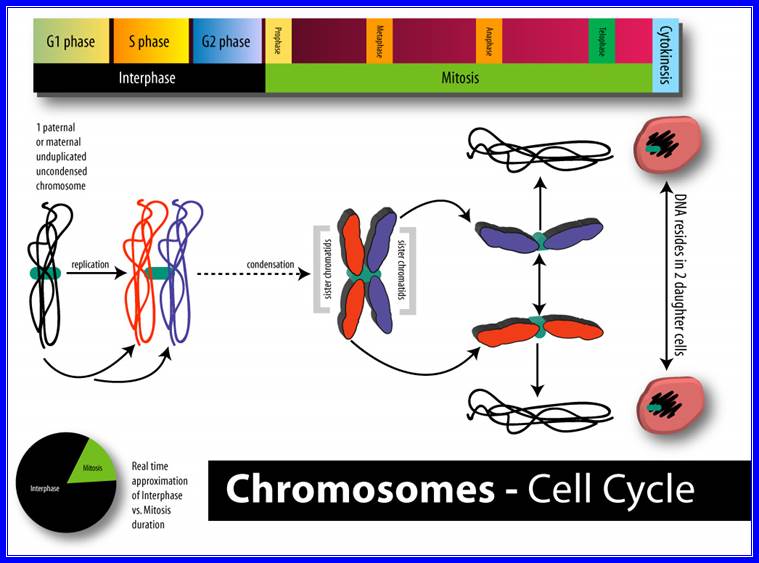
The process involves two important steps. The first is the division of nucleus (karyokinesis) and the second which normally follows karyokinesis is called cytokinesis. Further more depending upon the presence or absence of astral elements, it has been classified into astral type and anastral type. However, the whole process of cell cycle progresses sequentially through different stages, which merge with one another smoothly. Basing on the complex biochemical and physical changes, different stages have been recognized. The stages are interphase, prophase, metaphase, anaphase, telophase and finally cytokinesis.
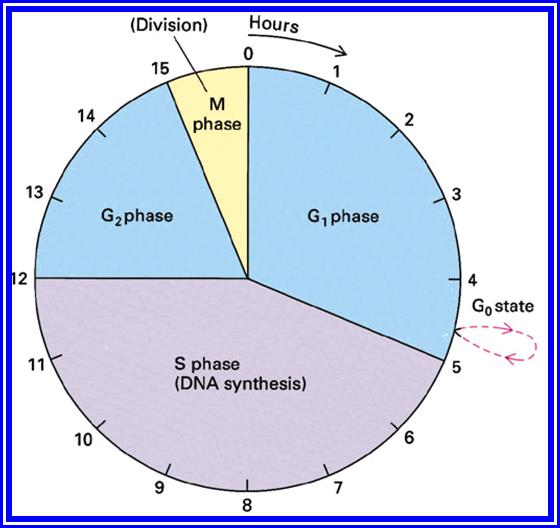
The time required for these different stages vary from cell type to cell type and organisms to organisms. Nevertheless, the interphase is the longest stage and the most variable. For example: in the root meristems of Vicia faba and Pisum sativum, the total time required for the whole cell cycle is about 24 hours, out of which interphase occupies about 23 hours and a half. On the other hand, prophase to telophase requires just about an hour and a half. On the contrary, in the corneal epithelial cells of rats, the whole process requires less than 30 minutes whereas the interphase takes up ~23 hours. Some of the embryonic cells take just 20 minutes complete one cell cycle.
Interphase: In yester years, interphase was considered as the resting stage, but in actuality, in dividing cells, it is one of the most active stages for it is the stage at which the cell prepares itself for the entire proceedings. Because of its long duration and varied biochemical activities this stage has been further sub-divided into phases like G1, S and G2 of which G1 phase is the most variable in duration. Intense biochemical activities take places at this stage and all precursors for DNA synthesis, histone synthesis are synthesized and mobilized. In multicellular cellular organisms where the cell division not required, the cell cycle gets arrested at this stage and undergoes a resting period called G° stage which ranges from few hours to years. At this stage, all cellular metabolism remains active to maintain the cell specific activities. Such a stage is called G0 phase. However, once the G1 stage is on, single stranded chromosomes move on to the next stage called ‘S’ phase. At this stage chromosomal DNA undergoes replication and produce two chromatin strands. During this process, the long DNA double helix unwinds and semi-conservative replication is initiated at several points simultaneously. Then the replication work progresses in both the directions till they meet the neighboring replicons. During replication, one of the daughter DNA molecules retains the parental histone core proteins and the other gets associated with the newly synthesized histone units to form a new chromatin thread. Thus two chromatin strands are formed in about seven hours of time, while G1 stage takes about 7-8 hours. Once the chromatin threads are duplicated, G2 phase is initiated. At this stage intense biochemical activities required for chromosomal contraction and development of mitotic apparatus take place. This state lasts for about 5-6 hours. All these activities ultimately result in the increase of nuclear size and now the cell is set to enter into a dramatic condensation and movement phase. It is important to note that through out this stage various types of RNAs are synthesized. Even as the chromatin DNA is undergoing replication, RNA synthesis continues.

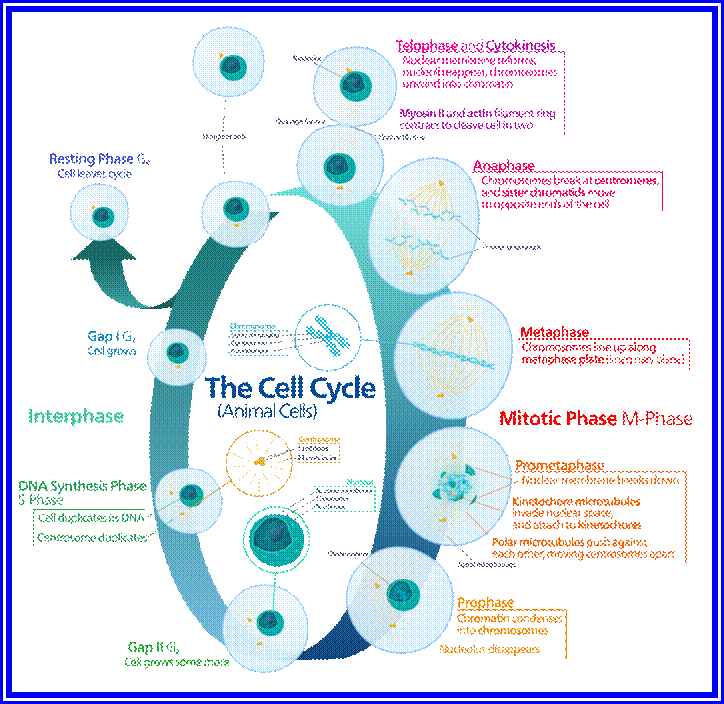
The phases (ordered counter-clockwise) of cell division (mitosis) and the cell cycle in animal cells.
Prophase:
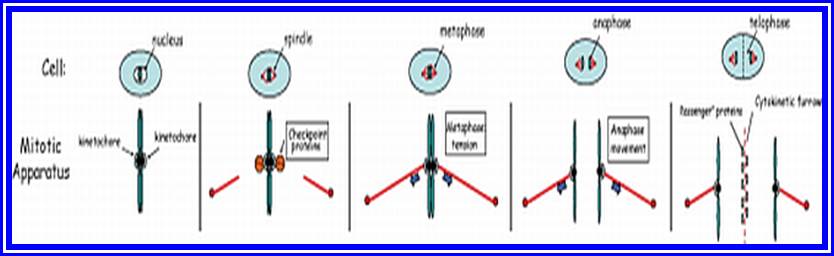
Prophase sets in as cell cycle factors operating in regulating cell division events are ready to go. The cell volume has reached a critical stage with all its components build up, the nucleus meanwhile also enlarges as the chromosomal DNA undergoes replication and the daughter chromatids fully form, the nuclear membrane starts dismemberment aided by lamin components as small membrane vesicles including pore complexes. Lamin phosphorylation, aided by CDK-Cyclins responsible for depolymerization of lamins and nuclear membranes just dismember. Appearance of mitotic apparatus and the dismemberment of nuclear membrane coincide.
Metaphase:
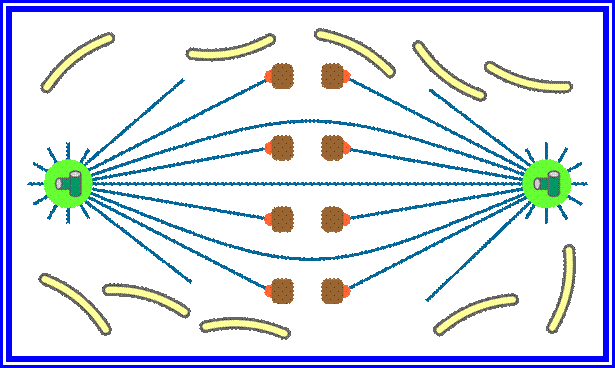
As the nuclear membrane disappears tractile fibers with continuous fibers appear in dome shaped form, which then called mitotic apparatus. Tractile fibers join or develop from kinetochore microtubular elements and extend. The other mitotic fibers are called continuous fibers emanate from poles. By contractile mode the tractile fibers bring all the chromosomes (which are divalent i.e. contain two sister chromatids), but the centromere is still intact and single. Interestingly, at CEN region, kinetochore complex organizes on either side of the CEN region, thus all mitotic chromosomes contain two kinetochore complexes; they with their tractile fibers are oriented toward their respective poles. It is at the end of the metaphase the CEN region appears to be split for the kinetochore bound by their respective tractile fibers are oriented towards their respective poles.
Anaphase:
Once duplicated chromosomes with their chromatids placed in the mid region of the cell or what is called equatorial plate, start moving towards their respective opposite poles.
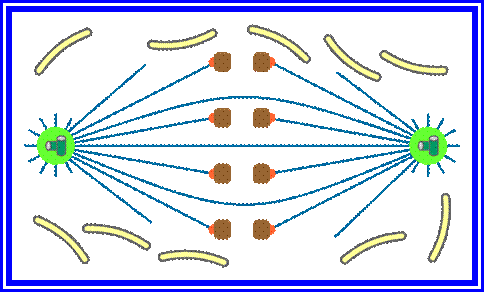
In astral type cells, the beginning of the prophase is indicated by the division of centrioles and the formation of radiating fibres all-round these daughter centrioles. As the nuclear volume increases, one of the astral points starts moving towards the other pole. Thus mitotic apparatus develops. Both in plant cells and animal cells tractile fibres which are associated with the kinetochore start depolymerizing at the poles. Thus chromosomes move along with the tactile fibres. Perhaps, simultaneously or at a little later stage, continuous fibres grow longer with the polymerization of tubulins from the polar ends. Thus they stretch the mitotic spindle and greatly aid in the movement of chromosomes. This process requires ATP as the energy source.
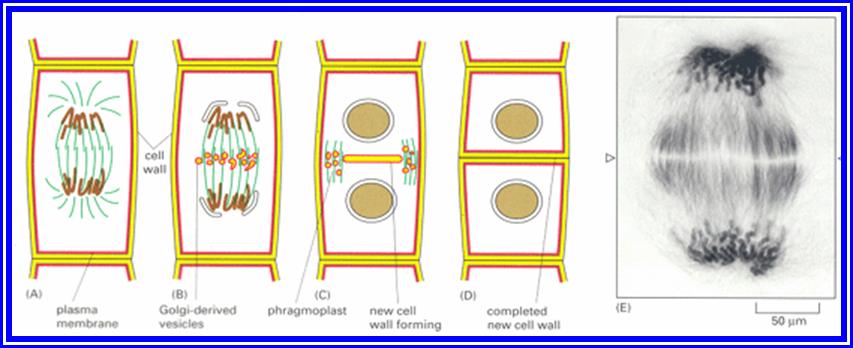
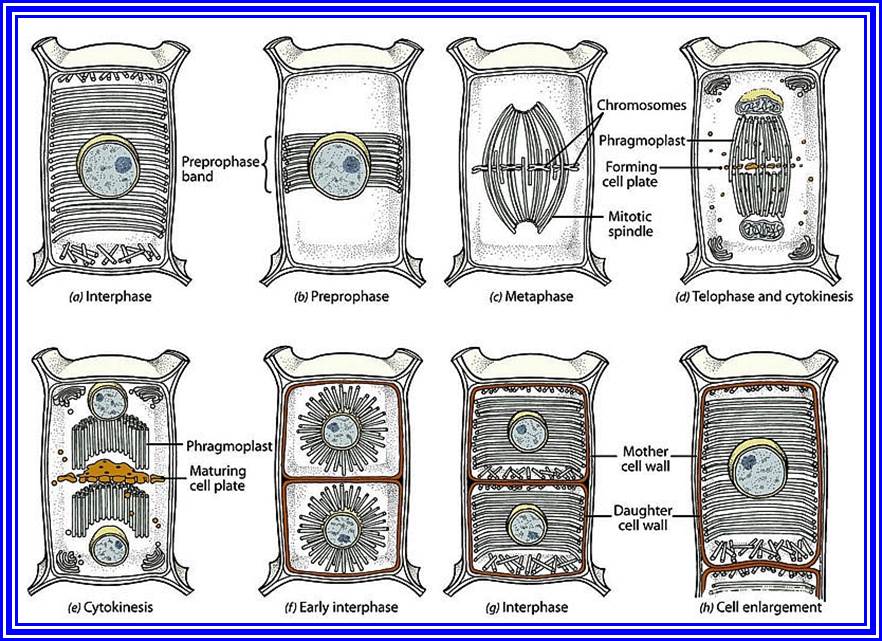
Cytokinesis means the division of the cytoplasm which results in the information of two daughter cells. cytokinesis of a typical plant cell differ from that of a typical animal cell. In a plant cell cytokinesis take place by means of cell plate formation. In the case by the end of anaphase a barrel shaped structure called phragmoplast is organised at the equatorial plane region of the cell. Vesicles formed by the near by Golgi complex enter into the centre of the phaplast and merge with each other.Pectin brought by these vesicles combine to form a fluid plate [cell plate] which extends and meets the mother cell wall. Fluid plate transforms into the middle lamellum.
Primary wall material is deposited on either side of the middle lamellum to complete the formation of cross wall and to finish off the cytokinesis.
In animals cells cytokinesis occurs at the end of telophase by constriction of the plasma membrane. The cell membrane invaginates almost in the middle of the cell to form a constriction which depends and finally the cell is divided into daughter cells.
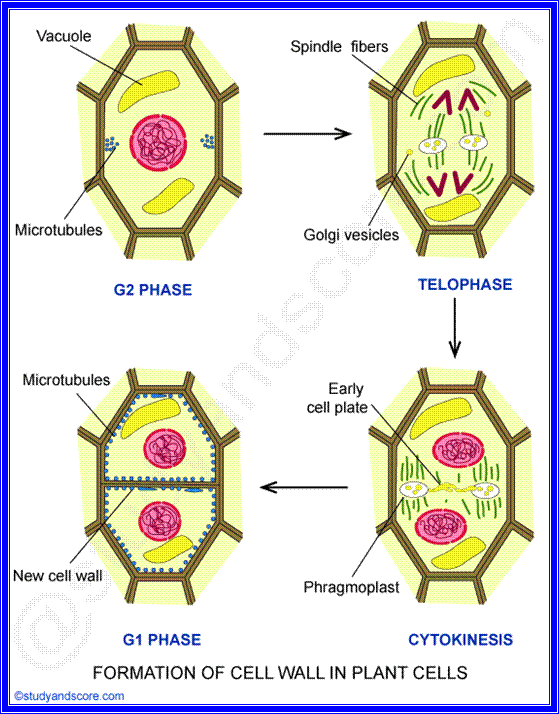
FORMATION OF CELL WALL
After nuclear division, when cytokinesis starts the cell wall is formed from the elements of Golgi complex and endoplasmic reticulum. The elements of golgi and endoplasmic reticulum align on the equatorial plate.
Middle lamella is formed by the deposition of calcium and magnesium pectates. Primary cell wall is secreted by protoplasm which lies inside middle lamella. Secondary cell wall is secreted by protoplasm which lied inside primary wall.
The following are the steps in the formation of the cell wall,
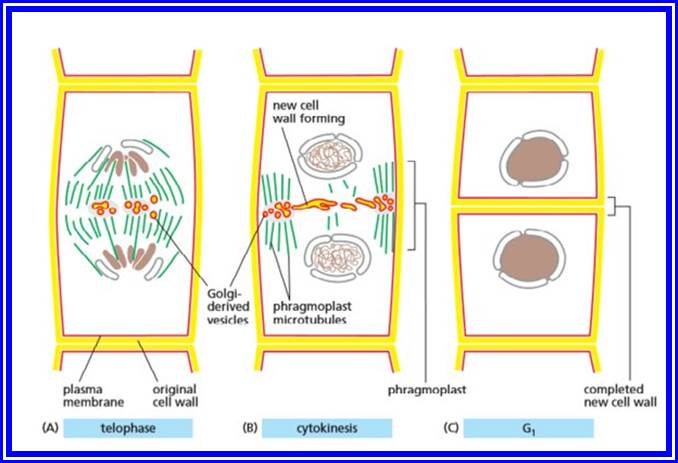
- During the telophase stage of mitosis, the phragmoplast widens and becomes barrel shaped. Then the cell plate begins to form inside the phragmoplast, on the equatorial plane
- In the area where the cell plate is formed, the fibres of the phragmoplast become indistinct and are restricted to the circumference of the cell plate.
· In plant cells, the division plane is predicted before mitosis by the location of a preprophase (PPB) band of microtubules and actin filaments in the cell cortex.
· The phragmoplast, a cytoskeletal structure that guides the formation of the new cell wall after mitosis, interacts with the cortical site formerly occupied by the preprophase band to position the new wall at that site. This interaction requires actin, and recent evidence indicates a possible role for myosins as well
· The plane of division for many plant cells can be predicted by their shapes. Cytoskeletal filaments radiating from the nucleus to the cortex may 'read' cell shape to determine the plane of division.
· In asymmetrically dividing cells, the plane of division is coordinated with cell polarity, which has been shown to depend on actin, the cell wall and secretion.
· In asymmetrically dividing cells of the maize leaf epidermis, Brick1 and Pangloss 1 are required for polarization of the mother cell and two Discordia genes are required for guidance of phragmoplasts to the former PPB site.
· The Tangled1 gene of maize is required for phragmoplast guidance in most or all leaf cells. Molecular analysis of the Tangled1 gene and protein indicates that TAN1 protein might participate in the orientation of cytoskeletal structures in dividing cells through an association with microtubules.
- When the cell plate is completely formed the phragmoplast disappears completely. At this stage thin lamellae are laid down by the daughter protoplasts on both the sides of the cell plate. The cell plate gradually undergoes changes to form the intercellular substances referred to as the middle lamella. Just under the middle lamella primary and secondary walls are deposited depending on the age of the cell.
Growth of Cell wall:
Just after formation, the cell wall is very thin and delicate. Later it grows by the deposition of the substances on the primary cell wall. The growth of cell wall occurs by two methods described as follows:
Intussusception: The growth that occurs by deposition of the cell wall material into the spaces present with in the cell wall is called as intussusception. As a result, the surface area of the cell wall increases.
Apposition: New cell wall materials are deposited on the surface of the existing cell wall in the form of thin layers. It results in the increase in the thickness of the existing cell wall. Apposition is also called accretion.
For various functional purposes, additional materials are deposited in the secondary cell wall. These depositions include:
- Lignin: Lignin is touch, glue-like substance deposited in the cells of woody plants. It helps in binding the wood fiber cells and also provides strength. Water and dissolved materials can pass through lignified walls. Deposition of lignin is called lignification.
- Cutin: It is wax like deposit found in the epidermal cells of the exposed surfaces.it makes the surface of the cells impermeable to water. Its deposition is called cutinization. Cutin deposition is very thick in xerophytic plants.
- Suberin: It is a fat like deposit found in the secondary wall of cork cells. It makes the cell wall impermeable to water. It also helps in checking the evaporation of water. Its deposition is called suberization.
- Minerals: Minerals like silica is deposited in the cell wall of grasses. Calcium oxalate and calcium carbonates are found in the cell walls of Acantharean, Cucurbitaceae and Moraceae.
- Mucilage: It is like carbohydrates which imbibe water and retains it for long duration. Mucilage is mainly found in seed coats and in aquatic plants
Sliding Theory: This theory envisages the presence of microfilaments associated with microtubules. The presence of Actin and Myosin units at the polar regions has been detected by antibodies raised against actin and myosin. Similar to that of muscular contraction, the microtubules of tractile fibres interact with acto-myosin like proteins found at the poles and slide over each other. Thus, the contraction of tactile fibres towards the pole is brought about. This process also requires ATP as the energy source.
Present concept: Even though both the above said theories are attractive, each of them has its own drawbacks. It is presumed that both the mechanisms may be operating simultaneously. To begin with, the sliding mechanism starts pulling the tactile fibres at the poles, at the same time the tactile fibres undergo depolymerization and continuous fibres get elongated by polymerization of tubulins. In fact, one finds tractile fibers and continuous fibers which running parallel to each other are inter connected with motor proteins and they play a role in pulling the chromosomes to their respective poles. Nevertheless, the knowledge about the molecular mechanisms of the organization of mitotic apparatus and the exact role in chromosomal movement and cytokinesis is far from clear.
Telophase:
In this stage, single stranded chromatids, that are pulled towards their respective poles start aggregating; simultaneously chromosomes start decondensation. Thus, the chromosomal strands become longer. At this stage transcription activity of chromosomal DNA begins. At the same time the nuclear membrane vesicles start appearing all-round the chromosomes and soon these membranous bits with pore complexes organize into nuclear envelope. Nuclear lamins play an important role is the reappearance of nuclear membranes. The relaxed chromatin attaches to nuclear matrix proteins at regions called MARS, which are specific sequence in chromosomal DNA that bind to proteins. As the chromosomal strands recoil and relax, the nucleolar DNA present in the region of secondary constriction loops out and nucleolar region begins to get organized. The nacked rRNA genes in clusters start transcribing precursor rRNAs. It is at the same time very many pre-rRNA processing sno RNAs and their associated proteins assemble in the nucleolus. A little later the incoming ribosomal structural proteins assemble on r.RNA and nucleolus gets organized. It is at this stage all the required Sn RNAs and their SnRNPs are produced for splicing events. It is important to note that chromosomes at this stage remain single stranded and fixed to the inner nuclear membranes. Soon one finds the development of cajal bodies and other nuclear speckles.
Cytokinesis:
Generally, karyokinesis leads to cytokinesis, but in certain organisms like plasmodia, siphonales members and others, cytokinesis does not follow karyokinesis and repeated nuclear divisions lead to multinucleate cells or coenocytic cells. Similarly, during early development of liquid endosperm in coconut fruits, nuclei divide repeatedly before cytokinesis sets in. The mechanism of cytokinesis in animal cells and plant cells vary; in the former case it is achieved by cleavage and in the latter, cytokinesis takes place by phragmoplasts formation.
Cytokinesis by cleavage:
In animal cells, cytokinesis sets in at late anaphase or early telophase. The appearance of dense materials at the equatorial region of mitotic apparatus is the first indication of cytokinesis.
Animal- cells mitotic division
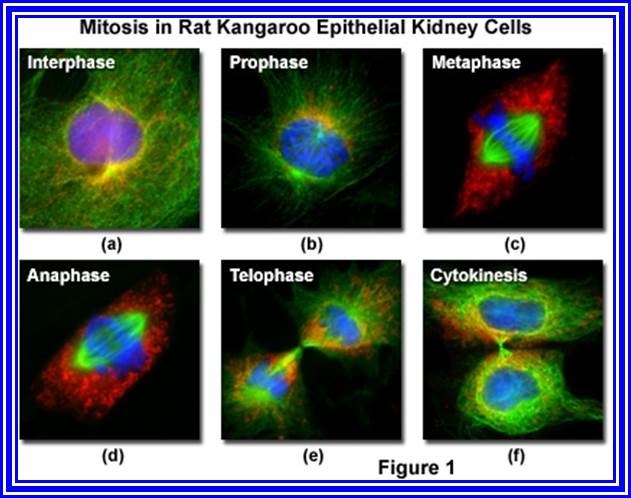
Great colored figures
However, during cleavage form of cytokinesis, at the equatorial region a ring of actin-myosin filaments appear in the cortical region of the cell. Using ATPase as the source of energy, these acto-myosin filaments interact. As a result, the associated protein filaments contract and the membrane to which these protein filaments are bound is drawn inwards all-round till the membranes fuse in the middle. Thus, the cells get separated. The exact mechanism of membrane contraction involving microtubules, actin and myosin and ATP is not clear. Nevertheless, the presence of these structures in and around the mitotic apparatus is known. Their involvement in the cleavage is just a presumption, of course with valid reasons.
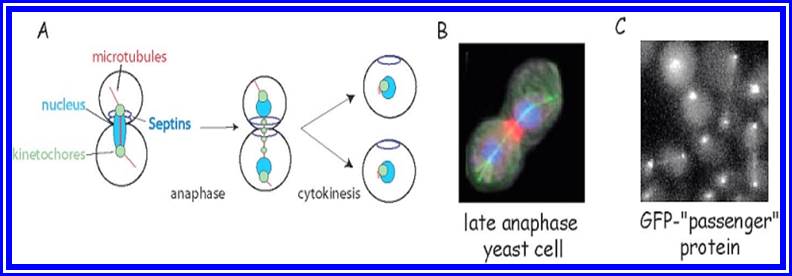
With the development of deep constriction some of the spindle fibres disappear due to disassembly of microtubules into tubulin monomers. Even after cleavage some remnants of microtubules that are found at the centriole region, also disappear at the end.
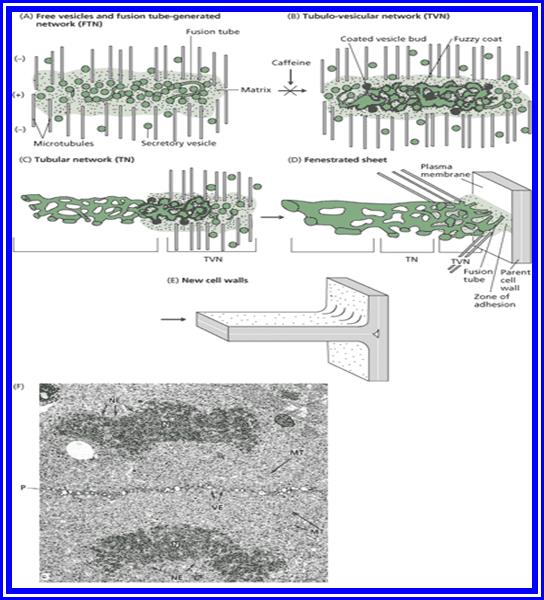
Cytokinesis by Phragmoplasts: Plant cells which do not possess centrioles, during cytokinesis, cells produce numerous membranous vesicles derived from Golgi complex and endoplasmic reticulum (SER). These vesicles appear at the inter-zonal region of the equatorial plate as well as at the microtubules found at this region. With time lapse, some more microtubules are added to the peripheral mitotic spindle. Thus, the mitotic apparatus appears to be bulged. Such a bulged structure of mitotic apparatus is called phragmoplasts. Later, the vesicles found in the equatorial region within the mitotic apparatus fuse with one another and form circular membranous cisternae, which gradually extend laterally and reach the phragmoplasts surface and the plasma membrane.
Cytoskeleton elements and their roles in Cell division
Finally, both the cisternae and phragmoplasts reach the lateral plasma membrane and fuse with it. Thus, the cytoplasm gets divided into two compartments by the membranous cisternae which act as the newly formed plasma membranes of the daughter cells. The space found between these membranes will be soon filled up by calcium pectate which acts as the middle lamella. Then the Golgi complex derived vesicles filled with cellulose fibres are directed with the help of microtubules towards the newly formed middle lamellae and a few cells wall, made up of cellulose fibres, is laid on either side of the middle wall. Thus, two daughter cells are produced.
Significance of Mitosis:
All multicellular organisms, as well as unicellular organisms use mitosis as a mechanism for multiplication of cells. During this process, chromosomes of parental cells duplicate and distribute equally to their daughter cells. Here the term ‘equally’ denotes both quantitative as well as qualitative. This process also helps in the growth of an organism. In many organisms where certain organs or cells are subjected to wear and fare, the cells are replaced continuously by mitosis, for example: in human beings there are about 2.5x1015; red blood cells and they have an average life of 420 days. In order to maintain the constancy of the blood cells, the body produces about 2.5 million new cells every second to compensate the loss, which appears to be incredible, but human body does it with mitosis.
Mitosis and Cancer;
Even though mitosis helps in the growth of an organism not only in the size but also in population, it is a highly related phenomenon. By mutational studies in yeast cells, as many as 38 or more steps have been identified to take part in mitosis, of which some are highly crucial in the progression of mitotic stages. If there are any mutations in the genome that control this process, cell division is completely inhibited or completely goes out of order or it may end up in an uncontrolled mitotic division. Under normal conditions, particularly in multicellular organisms, mitotically derived cells undergo differentiation and perform specific functions. Instead, in an uncontrolled process, cells undergo continuous multiplication by repeated mitosis. In these cases, the cell derivatives do not undergo any differentiation, but they divide and redivide endlessly. As a consequence of this, innumerable cells of the same kind are formed. Such a group of cells which are endowed with a potentiality to divide and redivide ceaselessly is called tumor cells and the disease thus produced is referred to as cancer. This can be induced by various carcinogenic agents like drugs, X-ray irradiations and even some viruses. Certain spontaneous mutations may also cause growth.
The analysis of cancer cells indicates that the rapid and uncontrolled cellular divisions are due to some changes in the regulatory chromosomal proteins called non-histones. Identification of such causative non-histones is very essential and important to cure the cancer disease.
In plants, however, callusing or callus formation is another example of uncontrolled, undifferentiated tumor formation. Nevertheless the callus formation is known to be controlled by certain phytohormones like Auxins. The special feature of these hormones is that at particular concentration, they induce tumor formation in plant cells, but at a different concentration with other hormones like cytokinins, they may induce differentiation of shoots or roots. The probable mechanism by which the hormones cause callus formation is again attributed to differential gene expressions or due to certain modifications of nonhistones proteins, which actually trigger off the cellular components to undergo such uncontrolled cell divisions.
Mitosis and cloning;
Development of a multicellular organism always begins with the zygote which is nothing but the product of syngamy. The zygote in turn undergoes repeated but controlled cell divisions which are followed by cell differentiation, where the cell derivatives develop into different types of cells which have their own characteristic structures and functions. The overall growth of an organism thus depends upon a controlled, determinate cell division and differentiation. The molecular basis of such cell differentiation is not clear, though certain differential gene expressions in E.coli, Drosophila and others have been very well studied.
Using the property of cell’s totipotency, where a single cell could be induced to develop into a complete organism, biologists have succeeded in the clonal propagation of plant in general and animals in specific cases. Normally, the production of off springs involves sexual reproduction, where two parents contribute the gene pool through gametes. Such off springs possess the mixture of genes from their parents. Instead, if diploid cells of one of the parents are induced to develop into an offspring, then such off springs are referred to as clones. Such clonal propagation is in vogue, particularly in plants, where the technique of tissue culture has been very well exploited.
In this process, a cell or a group of cells from any part of the plant body is explanted into a known solid or liquid agar based nutrient medium. If the medium is appropriate and balanced with the required phytohormones, the cell or cells explanted develop into callus, from which numerous embryos can be induced at will. Later, the embryos can be cultivated. This method has been successfully employed in cultivating horticultural plants, crop plants and also plants which are difficult to multiply by vegetative propagation or by sexual reproduction.
It is important to note, that this process has employed mitosis as the most important mechanism for cell multiplication. Nevertheless, this process of cell division is always followed or preceded by a regulated differentiation. Inspite of recent technical innovations, the molecular mechanism of differentiation is yet to be understood clearly. Drosophila and C. elegans have provided great insight into the molecular aspects of differentiation.
Unlike plants, clonal propagation of animals has been successful only in certain cases like frogs and rats. In these cases, diploid cells from the somatic tissues, rather than germinal cells, have been successfully used. Either by the transplantation of the somatic cell into the mother uterus, or by the transplantation of a nucleus taken out from the somatic cell into enucleated zygotic cell, complete animals have been grown in the laboratories. Such animals have the genome of only one parent and such off springs are called clones. Though clonal propagation of human beings has been attempted, the moral, social and ethical problems have deterred him from doing any further experiments. Nevertheless, with the time and change of attitude towards the fellow human beings, perhaps, one day he may resort to such clonal propagation of man to preserve himself.
Even though clonal propagation of higher plants and animals has its own implications as well as limitations, cloning of genes by Genetic Engineering techniques has been the craze of the day; its application in the welfare of fellow human beings is unlimited. Many genetic engineering industries have been set up in USA and other European countries. The trials and tribulations in developing this elegant but sophisticated technique are unsurpassed in the recent history of molecular biology. The pace of development in this field is phenomenal and it can be compared only to the space and computer technology. Biologists have already succeeded in cloning thousands of genes for insulin, growth hormone, interferon and many many more. The work is in progress to clone nitrogen fixing genes (nif genes) into eukaryotic plants. Hitherto, man has relayed on specific organisms as the source of gene products, unfortunately the labor, time and money spent to extract them was exorbitant. Added to this, the recovery was extremely poor. But the cloning techniques have made life easy and these products can be synthesized on a large scale, thus the cost of production as come down which is a great boon for common man.
MEIOSIS
Plants like ferns reproduce by spores and sweet peas by gametes. Higher animals produce gametes as a mode of reproduction. In all the above said cases and all other organisms, the plant or the animal body is diploid (2 n), such organisms resort to sexual reproduction by means of gamates. Sexual reproduction is just not restricted to higher system; this process is also prevalent
In bacteria.
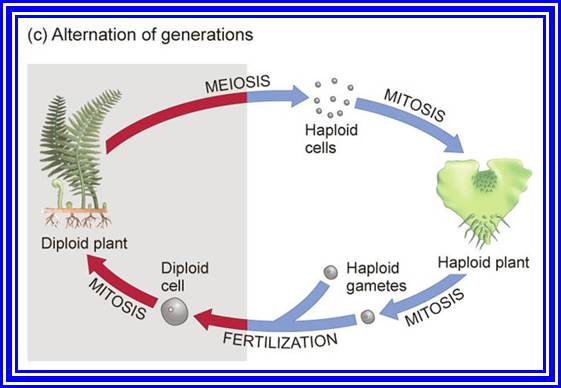
How mitosis and meiosis operate for the production of Humans and plants.
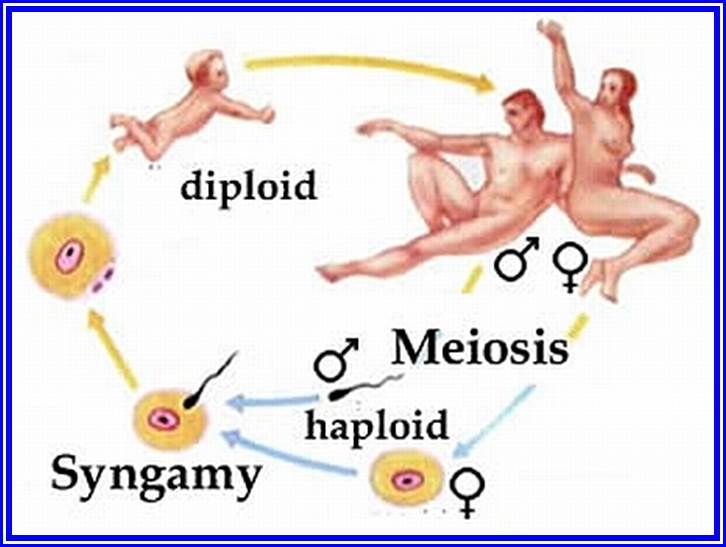
Meiosis is an important process for the production of gametes for the next generation.


Gregor John Mendel
Stages of mitosis
Meiosis.
The gametes like sperms and eggs fuse to produce a zygote which is the first cell generation of a new offspring. For example: human body is made up of diploid cells. The reproductive organs are also diploid. If the cells of these germ lines produce gametes of diploid nature, the fertilized product will be tetraploid. And in successive generations the ploidy level goes on increasing. But this does not happen; the off springs of the diploid parents will be always diploid. This is achieved by a remarkable process called Meiosis. The diploid reproductive cells produce haploid gametes by Meiosis, and such haploid gametes fuse to produce diploid off springs thus the diploid chromosomal number is maintained between parents and off springs by successive meiosis and fertilization. Thus, Meiosis is often considered as an antithesis for fertilizations.
Mechanism of Meiosis
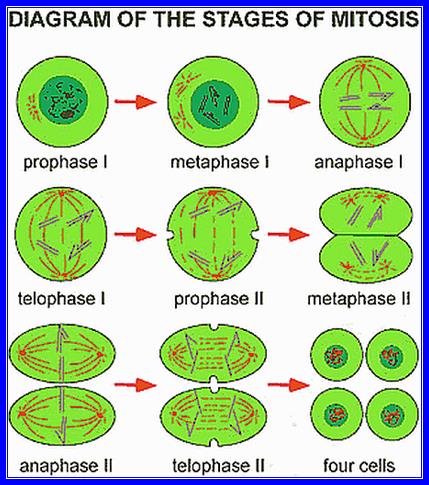
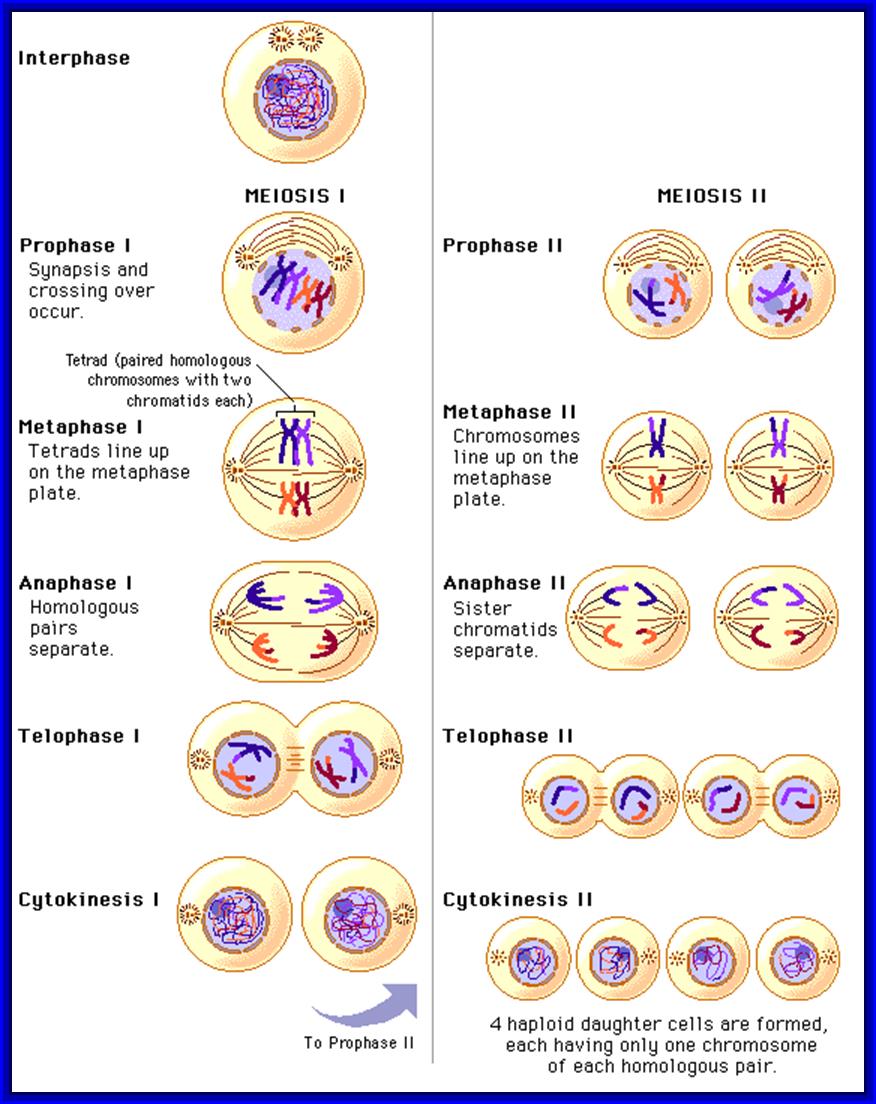
Meiosis unlike mitosis takes place in spore bearing or gamete producing cells. Most of the cells which are set to undergo meiotic division are quite large, distinct, and rich in cytoplasm and possess large nucleus with a conspicuous nucleolus. The cells show high rate of metabolic activity. The duration of meiosis varies from organism to organism and from few hours to many days, probably years. It takes place in two successive stages (1) Meiosis I or Reduction state (2) Meiosis II or Equational division. Each of these stages further, shows sub stages like Interphase, Prophase, Metaphase, Anaphase, Telophase and Cytokinesis.
Meiosis
Interphase: This stage is a preparatory stage for the subsequent events of karyokinesis. This, like mitosis, is also subdivided into G1, S & G2 stages.
G1 Stage: It occupies quite a large period of time at which cell prepares for the duplication of chromosomes. The necessary precursors like nucleotides, proteins and many required molecules are mobilized. As a consequence of this accumulation, the nuclear volume increases. The chromosomes found at this stage are extremely thin and they are not clearly visible except for certain heteropycnotic chromatin regions which appear as darkly stainable segments.
S-Stage: At this stage, hitherto single stranded chromosomes undergo duplication (through DNA replication, histone and nonhistones assembly) into double stranded chromosomes. Still the chromosomes are not clearly visible. However the nuclear volume further increases.
G2 Stage: This stage is comparatively of shorter duration and cell enters into prophase. The details of molecular events are not clearly known.
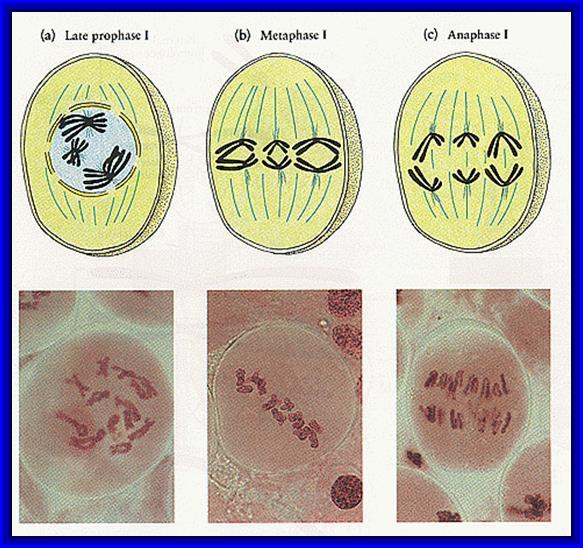
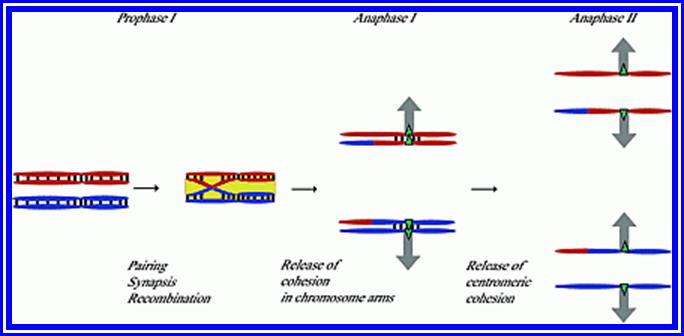
Prophase I: This stage is relatively longer and most complex; basing on the behavior of chromosomes and appearance, this stage has been further divided into sub stages: sequentially these are referred to as leptotene, zygotene, pachytene, diplotene and diakinesis. In all these stages, chromosomes continuously undergo condensation till metaphase.
Leptotene: At this stage, the most invisible chromosomes gradually become visible. This is due to condensation but still they appear to the single stranded structure, but are actually double stranded because of chromosomal duplication in the first interphase. Here, most of the chromosomes appear to be looped into horse-shoe shaped structures, where the chromosomal ends are found to be associated with the lamin region of the nuclear membrane. This association has been considered as very important for some proteins that are synthesized in the cytoplasm are transported and get associated with chromosomes through the nuclear membranes. This appearance of chromosomes has been referred to as “bouquet stage”, at this stage; chromosomes show fine granular or bead like structures called chromomeres. They were equated to genes by Bellings but later they were found to be nothing but coiled expressions of chromosomes.

Chromosomal features that express products
Zygotene: This is a stage which homologous chromosomes (derived from paternal and maternal side), start recognizing each other and initiate pairing. The pairing may be initiated at any point, i.e. terminal, middle or any where. However, once the pairing is initiated it proceeds like a Zipper. The process is so exact and precise; the pairing takes place– chromomere by chromomere and gene to gene. In any one of the homologous pair of chromosomes, if there is any non-homologous segments get paired the paring is rejected. If there are any segments of chromosomes which remain un-replicated, they replicate at this stage and the DNA that is replicated is called Z-DNA which is rich in GC content.
Synapsis.
Synapsis: The pairing of homologous chromosomes, though dramatic and fascinating to observe, the forces and factors that are responsible for recognizing each homologous chromosomes and mechanisms of movement of chromosomes, is yet to be completely resolved. Nevertheless, electron microscopic studies of some animal cells have revealed the presence of a ribbon shaped protein complex associated with the paired homologous chromosomes. This complex has been named as Synaptonemal Complex.
Synaptonemal Complex: This complex appears at the late leptonema or early zygonema and establishes fully at zygonema and pachynema. This structure starts disappearing at the end of diplonema. Synaptonemal complex is exclusively made up of protein units, arranged in between a pair of homologous chromosomes in the form of a paired ribbon. It consists of a pair of axials, a pair of lateral and a single central filament is the overlapping structure extended from the axial filaments. The space found between a pair of axial filaments; lateral filaments is 0.15µm to 2 µm. On the other side of the axial filaments, lateral filaments are present and these are associated with the homologous chromatin material. Some of the chromatin material penetrates through these protein complexes and open out into the central region as free DNA segments. These may consist of individual genes or a group of gene segments. The genetic material that is extruded from both the homologous chromosomes align parallel to each other, such that recombination may be brought about. Notwithstanding the above observation, the nature of the proteins and their functional mechanisms in pairing and recombination is solved to a great
extent. It is now known that the synaptonemal complex is made up of proteins, which not only recognize the homologous segments of chromosomes, but also bring about the pairing and recombination of genetic material between them. The proteins involved are complex, at least some of them bring chromosomes to be paired and some bring about condensation, and some bring about recombination.
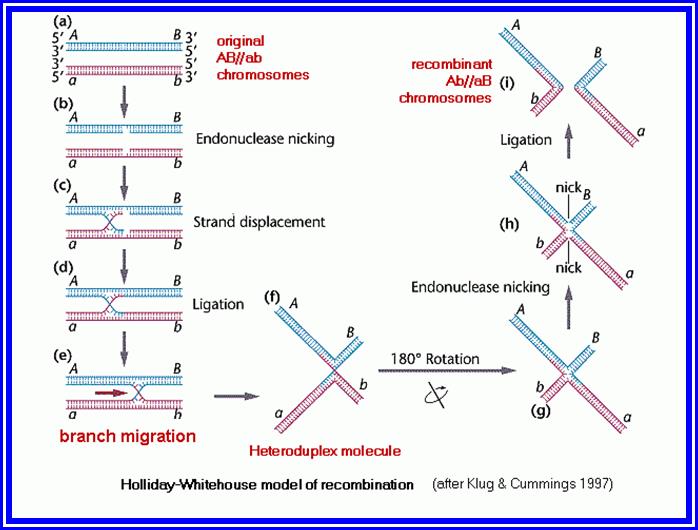
Pachytene: At this stage, if there are any parts of homologous chromosome unpaired, the paring will be completed. Then, at certain points, the chromosomal materials between homologous pairs exchange. This phenomenon is referred to as recombination or crossing over. This involves breakage and reunion between chromosomal strands. This involves homologous segments of chromosomes. Use of radio-active precursors have shown that the chromosomal material, get degraded and resynthesized. The enzymes that are involved in this process have been recognized as specific endonucleases, DNA polymerases, ligases and exonucleases.
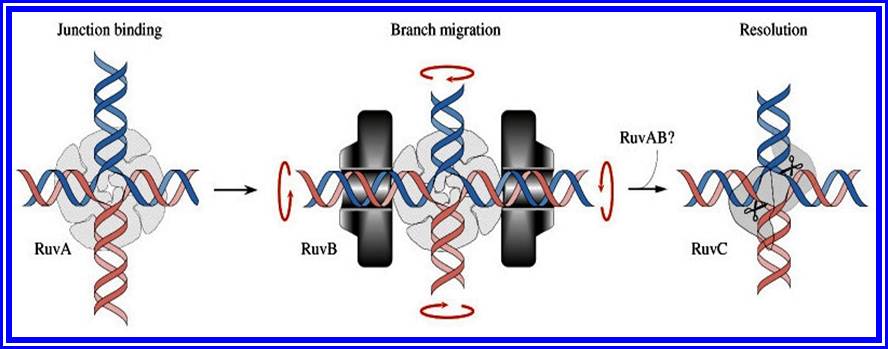
Mechanism of cross over: Basing on the microscopic observation of meiotic chromosomes, Darlington has opined that homologous chromosomes pair and coil relationally at zygotene under great strain. This coiling brings about great torsion and strain on chromonemal strands. Because of this, sister chromatids of both homologous chromosomes break at certain sites. This breakage releases the torsion and strain; the chromatids recoil and come to rest. If the broken ends of the non sister chromatids of two homologous chromosomes are brought near to each other or end to end, then they join, thus crossing over is brought about, resulting in genetic exchange between the parental chromosomes. Thus, crossing over is often summed up as the phenomenon of breakage and reunion between two non-sister strands of two homologous chromosomes. At the gross level this appears to be true. But break down and the reunion of chromosomes involve the break down of chromatin materials like proteins, DNA, and the rejoining involves the resynthesis and reassembly of the chromosomal proteins. Recombination is performed a variety of proteins such as Rec A, B and C.
In prokaryotic organism too, genetic recombination takes place at DNA level. It uses both homologous recombination as well as site specific recombination. It is not surprising to observe site specific recombination in viruses such as Lambda and HIV. It is not unusual to find in eukaryotes.
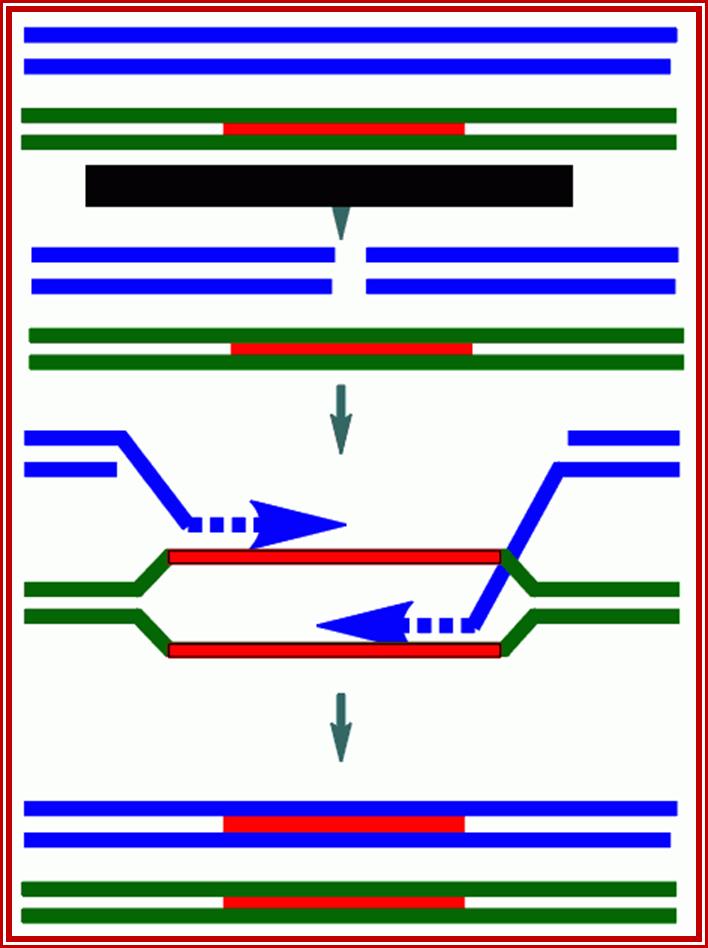
Molecular Mechanism of crossing over.
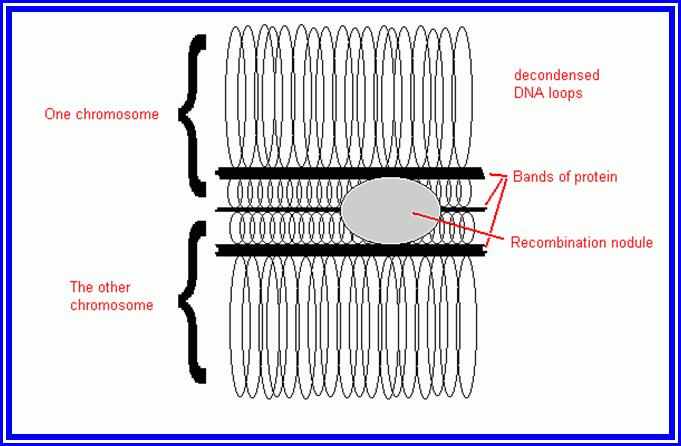
Molecular Mechanism of crossing over: When homologous chromosomes are brought together or paired by synaptonemal complex, some of the segments or chromosomal regions of homologous segments get disassociated with chromosomal proteins and the DNA strands from the apposing chromosomes loop out with the central region found between two axial filaments of SC. Synaptonemal complex consists of axial filaments, and lateral filaments, it also contains nodules which are endowed with recombination enzymes.
At various points, all along the length of chromosomes start aligning with each other so that the homologous DNA segments are brought near as well as parallel to each other.
The paring of homologous chromosomes is brought by a set of DNA binding proteins called adhesins. And contraction is brought by condensins. Then certain endonucleases found in synaptonemal complex cut open the DNA strands as shown in the figure.

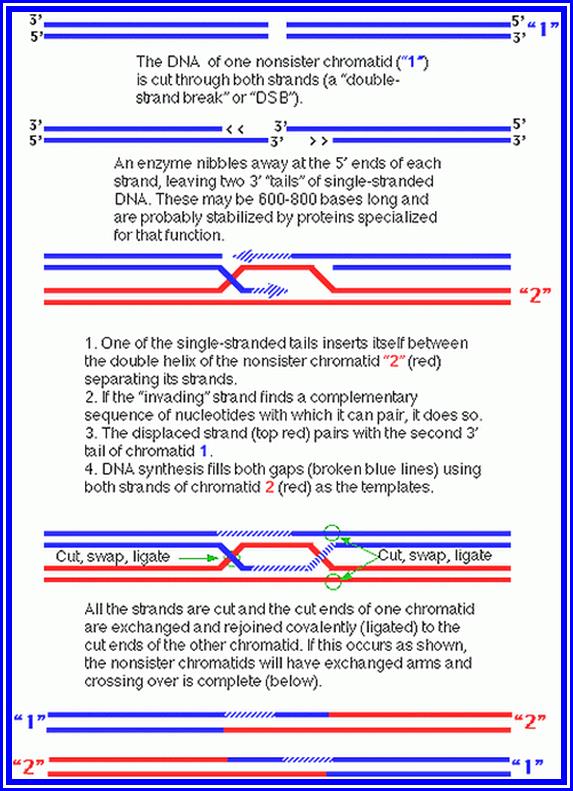
Crossing over between chromosomal pairs in Meiosis.
As a result, the DNA segments break open with long sticky ends of DNA belonging to homologous pair of chromosomes have complementary nucleotide sequences. This leads to the cross pairing of the single stranded DNA ends. This pairing may result either in the overlapping with some part of the segment sticking out or certain gaps may be left out. In the former condition exonucleases cut out the extra DNA segment and in the latter case, gaps are filled up by DNA polymerases. DNA synthesized at the time of genetic recombination is called P.DNA. However, the strands are finally sealed by DNA ligases. Thus, crossing over or recombination is brought about. Such recombination events, particularly in prokaryotic organisms like bacteria have been observed through electron microscope. But in Eukaryotic organisms the visibility, under TEM is very clear and distinct. However, indirect studies like the incorporation of radioactive DNA precursors into meiotic chromosomes, have added more evidence to theory, which is now a fact. Further more, the presence of dense nodules of 100 nm size found in the central component of SC have been assumed to be recombination nodules, involved in genetic exchange.
Diplonema: The duration of this stage varies from few hours to many years. For example, in human females, oocytes develop in the germ line, when the human embryo is still 5 months old. The oocytes start meiosis and reach up to diplonema.

At this stage, meiosis gets arrested till the females reach the puberty then the oocytes one by one complete meiosis and eggs are released periodically till the age of 50 years.
In this stage of diplonema, the pairs of homologous chromosomes start moving away from each other, probably because of the disappearance of synaptonemal complex. As the chromosomes repel at each, some parts of chromosomal segments are still held at the regions of chromosomal exchange, which show up as X configurations and they are called chiasmata. As the chromosomes further open out, the chiasma starts slipping away along the chromosomal strands. This phenomenon is called Terminalization.
Diakinesis: At this stage chromosomes complete the terminalization and whatever synaptonemal complex that has remained, disappears completely. Meanwhile the chromosomes reach maximum condensation.
The disappearance of nuclear membrane and nucleolus marks the end of Diakinesis and Prophase.
Metaphase I: With the disappearance of nuclear membrane, mitotic apparatus appears. The tactile fibres get attached to the kinetochore regions of the centromere of the double stranded chromosomes. Unlike mitotic metaphase chromosomes at this stage, each of the homologous chromosomes will have only one kinetochore, though centromere appears to be double. The tractile fibers drag the respective homologous chromosomes and each of the chromosomes will have only one kinetochore, though centromere appears to be double. The tractile fibers drag the respective homologous chromosomes on the metaphasic plate or equatorial plate and orient the homologous chromosomes in such a way, one of the homologous chromosomes is directed towards one pole and the other is to the opposite pole. Thus a pair of homologous chromosomes found on the equatorial plate will have one group of tactile fibres emanating from one side of the centromere of one set of chromosomes and another group of tractile fibers from the other set of chromosomes, which is directed to opposite poles. This particular orientation is remarkable, for it is this that ensures reduction division. Furthermore, the centromeres of the metaphasic chromosome do not divide and double strands of each homologous chromosome remain intact.
Another interesting, as well as important aspect is that during the orientation of homologous pair in the equatorial region, the maternal and paternal chromosomes are at random. This results in the random and impendent assortment of genes.
Anaphase: The respective homologous chromosomes are pulled to their respective poles. During this movement chromosomes undergo decondensation slightly. The mechanism of chromosomal movement is same as that of mitotic Anaphase movement.
Telophase: All the chromosomes that move towards their respective poles are pulled together and chromosomes undergo further decondensation resulting in the formation of chromatin network.
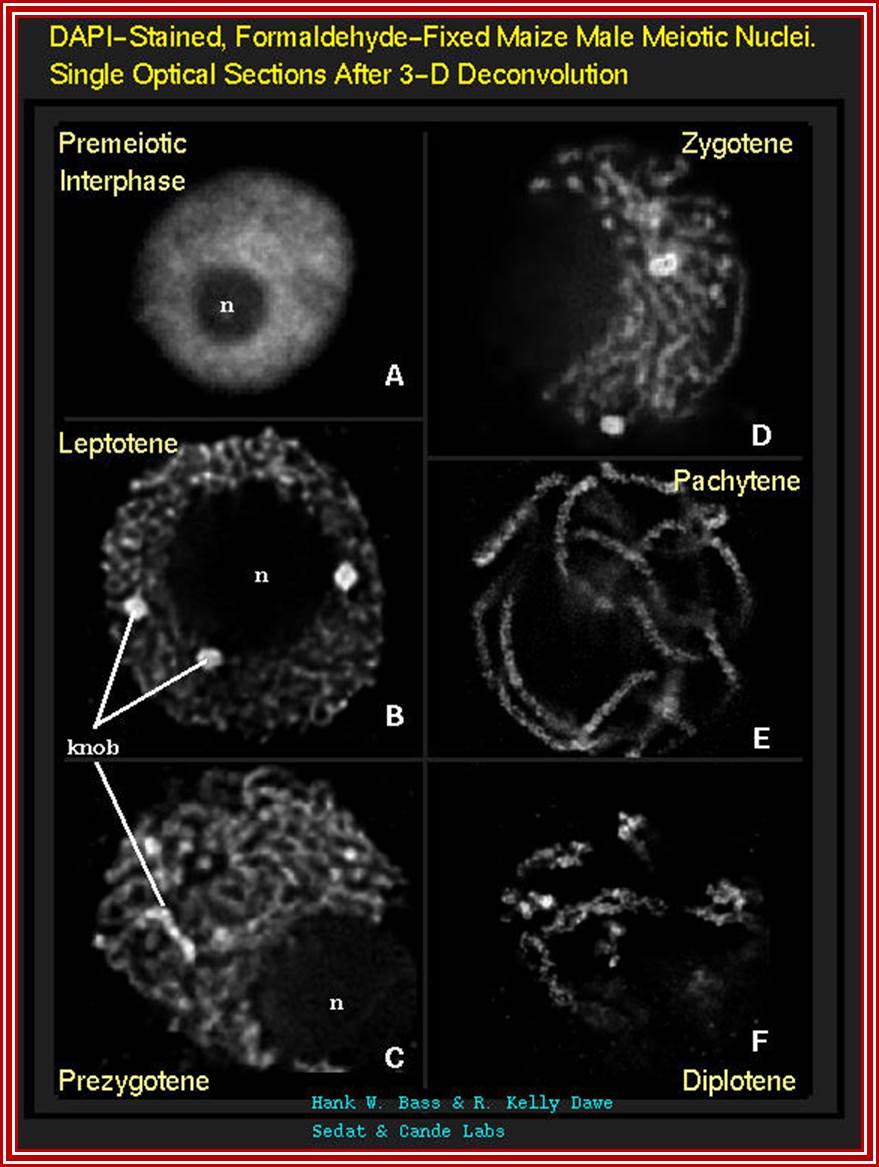
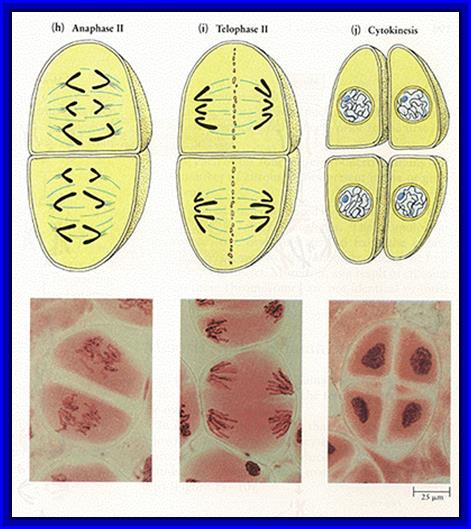
Micrographic pictures of chromosomal appearance at different stages.
Interphase II and Prophase II: The interphase is the shortest stage. Some times the meiotic cells skip the interphase and they directly move on the metaphasic stage.
Metaphase II: The mitotic apparatus appears at this stage with a different polarity. Chromosomes are brought to the equatorial region and the chromosomes are oriented in such a way that the tactile fibres originate from Kinetochore that develops on either side of the centromere. The tactile fibres are oriented in the opposite direction. The centromeric region divides into two, which frees individual chromatids free.

Anaphase II & Telophase II: The single stranded chromosomes are now pulled to their respective poles. During these stages, chromosomes undergo further decondensation. Finally, all the chromosomes in their respective poles get organized into a net work of chromatin. Later nuclear membranes and their nucleoli appear.
Thus, starting from the single diploid nucleus, four nuclei with haploid chromosomal sets are formed.
Cytokinesis: In the case of animal cells the cytokinesis is by cleavage, on the contrary phragmoplasts are formed in plant cells. However, the cytokinesis always takes place after completion of karyokinesis. This may be successive or simultaneous. The four cells they derived from meiotic division are called tetrads. These may further develop either into haploid spores or haploid gametes, which again depends upon the organism and the stage of life cycle.
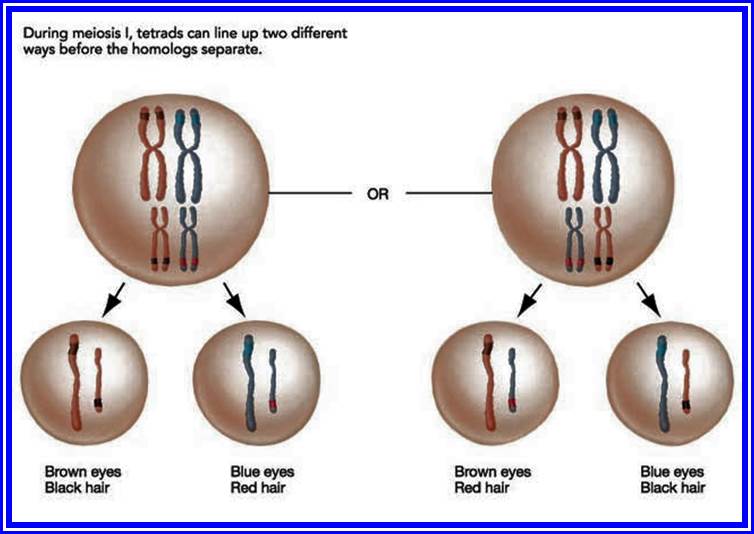


Importance of Meiosis:
1. It acts as an antithesis for fertilization. Genetic recombination takes place between parental homologous chromosomes.
2. As parental homologous chromosomes arrange randomly at first metaphasic plate, chromosomes segregate equal but independently.
3. Genetic recombination and random segregation result in the production of gametes with variable genotype.
4. Variable genotypes help in bringing about variation result in the production of gametes with variable genotype.
This variation plays a significant role in evolution.